Articulo original
Original article
35 (2) | Jul - Dic, 2025
Facultad de Ciencias Exactas y Naturales y Agrimensura (UNNE)
Open Access: https://revistas.unne.edu.ar/index.php/fce
E-mail: revistafacena@exa.unne.edu.ar
Análisis paleopalinológico, litofacial y paleoambiental de dos localidades de la Formación Palo Pintado (Mioceno Tardío), Salta, Argentina
Paleopalynological, lithofacial and paleoenvironmental analysis of two localities of the Palo Pintado Formation (Late Miocene), Salta, Argentina
Horn, Maricel Yanina * ¹ & Galli, Claudia Inés 2
1. Centro de Ecología Aplicada del Litoral (CECOAL, CONICET-UNNE). Corrientes (Capital), Argentina.
2. Centro de Estudios Geológicos Andinos (CEGA-CONICET), Facultad de Ciencias Naturales, Universidad Nacional de Salta, Salta (Capital), Argentina.
* Autor de correspodencia: yaninahorn@yahoo.com
Recibido/Received: 12 de Marzo, 2025 | Aceptado/Acepted: 27 de Septiembre, 2025 | Publicado/Published: 19 de diciembre, 2025.
Como citar este artículo: Horn, M. Y. & Galli, C. I. (2025). Análisis paleopalinológico, litofacial y paleoambiental de dos localidades de la Formación Palo Pintado (Mioceno Tardío), Salta, Argentina. Revista FACENA 35(2), 120-148. Doi: https://doi.org/10.30972/fac.3528179
Resumen: El presente trabajo se enfoca en el estudio de los palinomorfos recuperados de muestras sedimentarias de las localidades Quebrada El Estanque y Río Calchaquí de la Formación Palo Pintado (Mioceno Tardío) Salta, Argentina. Se determinaron paleocomunidades, y establecieron relaciones entre litofacies y paleoambientes de depósito. La Formación Palo Pintado está constituída por areniscas y pelitas, mientras que los conglomerados están subordinados. El conjunto muestra una disposición en ciclos que han sido interpretados como sistemas fluviales de baja jerarquía y baja movilidad. En la Quebrada El Estanque se hallaron veintiséis especies de palinomorfos, que habrían integrado las paleocomunidades acuática, palustre, xerófila y boscosa de altura. En cambio, en Río Calchaquí, se hallaron diez especies de palinomorfos que habrían conformado las paleocomunidades bosque higrófilo, xerófila y boscosa de altura. En Quebrada El Estanque el ambiente de sedimentación correspondería a llanuras inundables en las que podrían haberse formado lagunas por desbordes de los ríos. En las orillas habría tenido lugar el subambiente palustre y en la zona litoral se habría localizado la vegetación palustre herbácea, mientras que las especies xerófilas se habrían encontrado en zonas más alejadas. En Río Calchaquí, los sedimentos corresponden a acumulaciones de canal o intracanal con escaso desarrollo de llanuras inundables. La vegetación higrófila se habría desarrollado en las cercanías de los ríos, en tanto la vegetación xerófila en ambientes abiertos. El clima habría sido cálido húmedo con estacionalidad seca.
Palabras claves: Palinomorfos; Paleocomunidades; Formación Palo Pintado; Cenozoico.
Abstract: The present work focuses on the study of the palynomorphs recovered from sedimentary samples from the Quebrada El Estanque and Río Calchaquí localities of the Palo Pintado Formation (Late Miocene) Salta, Argentina. Paleocommunities were determined, and relationships were established between lithofacies and depositional paleoenvironments. The Palo Pintado Formation is composed of sandstones and mudstone, while conglomerates are subordinate. The deposits show an arrangement in cycles that have been interpreted as river systems of low hierarchy and low mobility. Twenty-six species of palynomorphs were found in the Quebrada El Estanque, which would have been part of the aquatic, marsh, xerophilous and high-altitude forest paleocommunities. On the other hand, in Río Calchaquí, ten species of palynomorphs were found that would have integrated the paleocommunities of hygrophilous, xerophilous and high-altitude forest. In Quebrada El Estanque the sedimentation environment would correspond to floodplains in which lagoons could have formed due to river overflows. The marsh subenvironment would have taken place on the shores and the herbaceous marsh vegetation would have been in the coastal area, while the xerophilous species would have been found in more distant areas. In the Río Calchaquí, the sediments correspond to channel or intrachannel accumulations with little development of floodplains. Hygrophilous vegetation would have developed in the vicinity of river edges while xerophilous vegetation in open environments. The climate would have been warm, humid with dry seasonality.
Keywords: Palynomorphs; Paleocommunities; Palo Pintado Formation; Cenozoic.
Introducción
Los depósitos sedimentarios cenozoicos de la Formación Palo Pintado, se encuentran ubicados en la zona sur del Valle Calchaquí, a unos 200 km al suroeste de la ciudad de Salta (Figura 1). A lo largo de los años sus afloramientos en distintas localidades fosilíferas (como Quebrada El Estanque, Quebrada de Alfredo, Quebrada el Horno, Quebrada Rincón del Horno, Quebrada de Salta, Santa Rosa, La Viña, Peñas Blanca entre otras) han sido estudiados por diferentes grupos de paleontólogos y geólogos. El Dr. Rafael Herbst y la Dra. Luisa, M. Anzótegui desde la década del 80 hasta el año 2012 aproximadamente, recorrieron la Formación con el objetivo de hallar impresiones de hojas, tallos, semillas, frutos, leños y tomar muestras sedimentarias para la recuperación de palinomorfos. Los resultados de sus investigaciones se plasmaron en publicaciones científicas en coautorías con otros investigadores o becarios (Por ejemplo: Acevedo et al., 1997; Anzótegui 1998, 2006; Anzótegui y Cuadrado 1996; Anzótegui y Horn 2011 a y b; Anzótegui et al., 2017, 2019; Barreda et al., 2007; Galli et al., 2011; Herbst et al., 1987, 2000; Horn et al., 2011; Robledo et al., 2015, 2020; Starck y Anzótegui 2001). Además de la coautoría de algunos trabajos compartimos con ellos el viaje de campaña donde se tomaron las muestras de la localidad Quebrada El Estanque. Particularmente, la Dra. Anzótegui (Yuyi) fue Directora de la Tesis Doctoral de Horn (2014) para la cual se estudiaron las dos localidades cuyos resultados se presentan a continuación. Por estos motivos redactamos este artículo especialmente para ella, a modo de homenaje y reconocimiento a su importante aporte al conocimiento de la paleoflora neógena del Norte de Argentina.
El registro fósil de esta Unidad también cuenta con restos de invertebrados dulceacuícolas (Morton 1992; Morton y Herbst 2012), escamas y vértebras de peces, placas y huesos de otros vertebrados (Armella et al., 2016; Bona et al., 2014; Candela et al., 2021; Robledo et al., 2023; Zimic et al., 2018; Zurita et al., 2024).
El presente trabajo tiene como objetivos contribuir al conocimiento palinológico de la Formación Palo Pintado asociado a su paleoambiente depositacional, para lo cual se centró en: a) determinar las diferentes especies de palinomorfos (granos de polen, esporas, colonias/cigósporas de algas, quistes de algas esporas de hongos, briófitos y helechos) hallados en las localidades Quebrada El Estanque y Río Calchaquí y reconocer las diferentes paleocomunidades que habrían integrado, b) relacionarlas con el paleoambiente sedimentario que surge del análisis de las litofacies y estructuras sedimentarias, c) aportar información a la existente sobre la paleoflora del Neógeno de Argentina y d) desde el punto de vista taxonómico contribuir a la biodiversidad de la región.
Marco geológico
El área de distribución de la Formación Palo Pintado ocupa una franja de sentido Norte-Sur en el Valle Calchaquí, extendiéndose desde el paralelo de 25° 30’ a 25° 50’ latitud sur (Díaz y Malizzia, 1983). Integra el Grupo Payogastilla (Díaz y Malizzia, 1983), de base a techo, conformado por las Formaciones Los Colorados, Angastaco, Palo Pintado y San Felipe (Díaz et al., 1987). El área donde afloran las localidades estudiadas se encuentra ubicada en la zona sur del Valle Calchaquí (entre San Rafael, al norte de San Carlos y Angastaco), entre los 25º41’01”S-66º07’55”W y los 25º40’59”S-66º05’49”W, aproximadamente a 200 km de la ciudad de Salta (Galli et al., 2011) (Figura 1). Las sedimentitas de la Formación Palo Pintado son fácilmente reconocibles por su color pardo amarillento y verdoso, con una disposición en ciclos grano y estratodecrecientes de conglomerados soportados por matriz, areniscas cuarzosas, que culminan con niveles de pelitas verdes.
A partir de los 10 Ma se inicia la acumulación de las sedimentitas que conforman la Formación Palo Pintado (1000 a 1400 m de espesor), hasta los 5 Ma. En este intervalo temporal se instaura un sistema fluvial, bajo condiciones climáticas tropicales y húmedas, conformando un sistema fluvial sinuoso areno-gravoso con desarrollo de lagunas y pantanos (Galli y Reynolds, 2012; Galli et al., 2011, 2017).
En base a las características de la arquitectura fluvial y sus litofacies asociadas se distingue un sistema fluvial con depósitos desarrollados dentro del canal o intracanal y de overbank. Las acumulaciones intracanal están conformadas por depósitos de barras gravosas (elementos GB) y formas de lecho que se relacionan con barras transversas y sand wave por acreción vertical y corriente abajo (elemento SB). Mientras que los depósitos de overbank están representados por: a. barras de acreción lateral, en donde la superposición lateral y vertical representa repetidos procesos de erosión y acreción lateral en la llanura de inundación (elemento LA); b. canales pequeños producto de la erosión en los bordes del canal principal durante eventos de inundación, los cuales son asignados a crevasse channel (elemento CS) y c. desarrollo de una amplia llanura de inundación (elemento FF) (Galli et al., 2011) (Figura 3).
Los resultados de la presencia de minerales de arcilla en el subambiente de llanura de inundación, como illita, esmectita y caolinita indican que han sido generadas por hidrólisis en un clima templado-húmedo (Galli et al., 2010, 2011 y 2014 a y b; Starck y Anzótegui 2001). La formación de estos niveles arcillosos para el tiempo de depósito de la Formación Palo Pintado evidencian un cambio climático importante entre los 10 y 5 Ma en el sistema fluvial sinuoso areno-gravoso, que no se registran en los niveles más antiguos de la misma cuenca de depósito. La presencia de Caiman cf. latirostris soporta la hipótesis de las condiciones climáticas tropicales para este intervalo de tiempo (Bona et al., 2014).
La localidad Río Calchaquí se encuentra sobre la Ruta Nacional Nº 40 a metros del puente que cruza el Río Calchaquí (en las cercanías de la localidad de Santa Rosa, (Figura 1). Estratigráficamente se encuentra representada la sección inferior de la Formación Palo Pintado, a partir del límite con la Formación Angastaco. Se realizó un perfil de 200 m de espesor, integrado desde la base a techo por niveles de areniscas medianas y gruesas, intercaladas con capas de conglomerado fino, mediano y grueso; pelitas verdes y marrones. En la sección basal del perfil se encuentra un nivel de toba.
La localidad Quebrada El Estanque, se encuentra ubicada entre las localidades de Angastaco y Santa Rosa (25°40’48.87”S- 66° 4’37.68”O, Figura 1), se exhibe en la sección medio-superior de la citada unidad. Se realizó un perfil de 362 m de espesor, conformado por niveles pelíticos (amarronados, algunos verdosos), alternados con potentes areniscas (duras, amarillentas), intercaladas con delgados niveles conglomerádicos, los cuales forman ciclos de origen lacustre. Se observó también en algunos de ellos la presencia de restos fósiles de vegetales (tallos, hojas), escamas de peces y bivalvos.
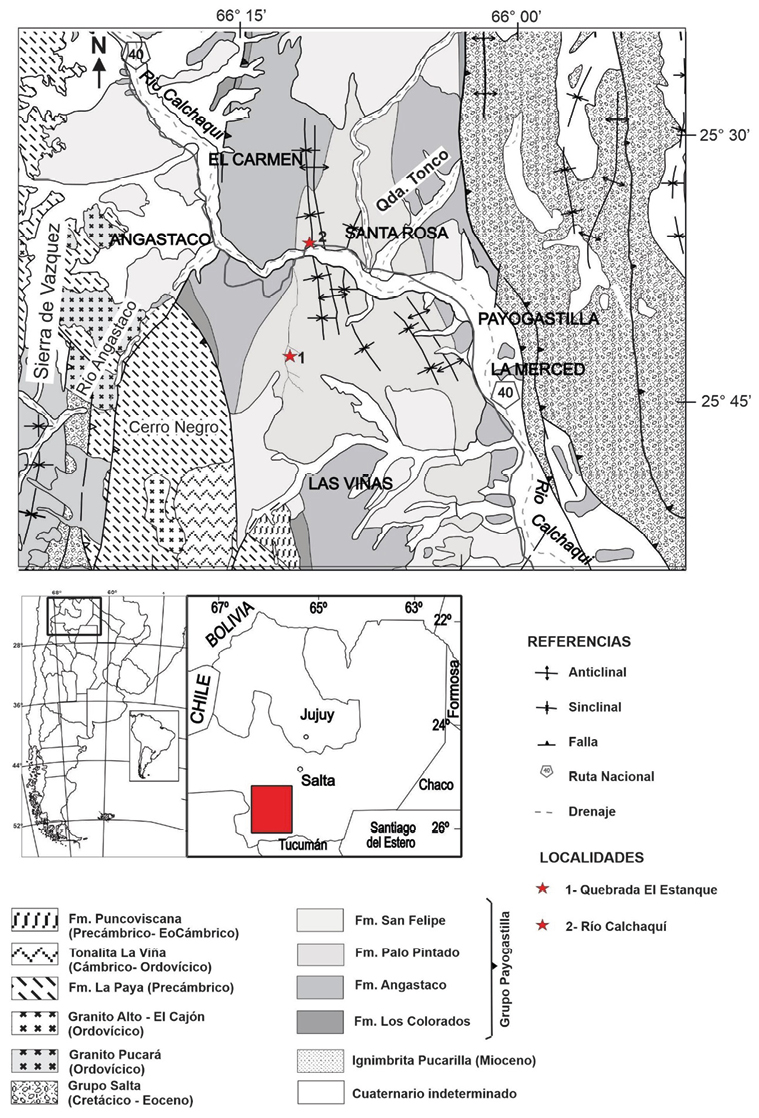
Figura 1. Mapa Geológico y de ubicación geográfica de las localidades fosilíferas (véase recuadro al SO de la provincia de Salta). Tomado de Hongn y Seggiaro (2001); Salfity y Monaldi (2006).
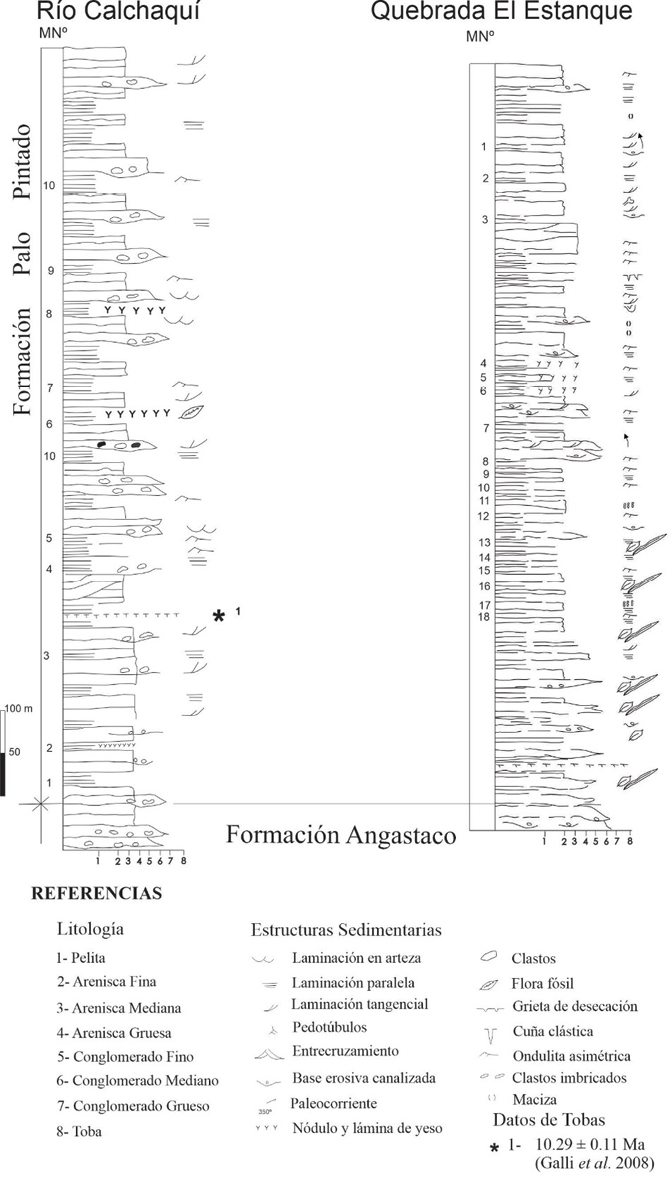
Figura 2. Columnas estratigráficas de las localidades Río Calchaquí y Quebrada El Estanque, señalando los niveles estudiados. En la segunda columna el orden de los números de muestras (MN°) se encuentra invertido en relación a la primera, en correlación con el orden de los niveles sedimentarios en el diagrama polínico.
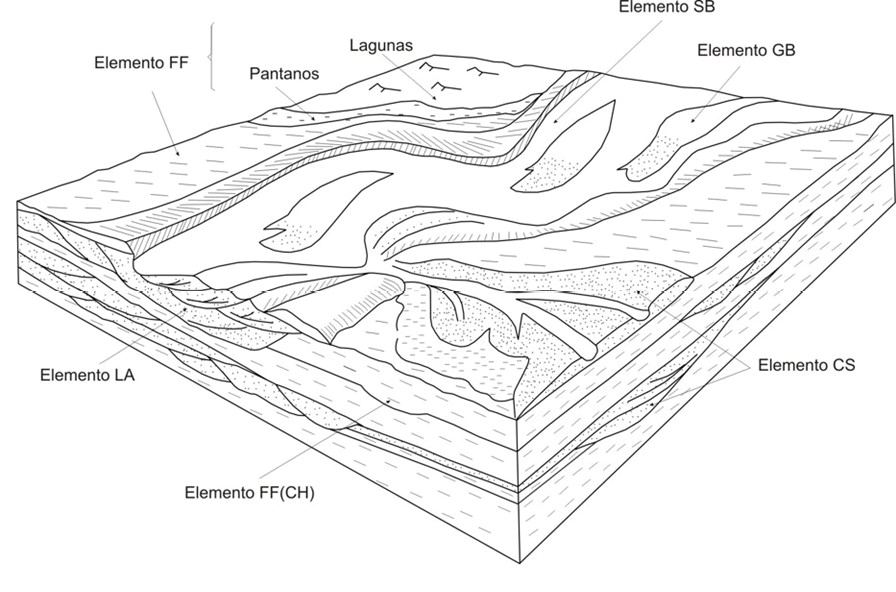
Figura 3. Diagrama esquemático de la distribución de los elementos arquitecturales interpretados para la Formación Palo Pintado. Elemento SB (barras transversas y sand wave), elemento CS (crevasse splay), elemento LA (depósitos de acreción lateral), elemento FF(CH) (relleno de canales abandonados), elemento GB (barras gravosas), elemento FF (llanura de inundación) (Tomado de Galli et al., 2011).
materiales y métodos
Se tomaron un total de 74 muestras sedimentarias entre las dos localidades (64 Quebrada El Estanque y 10 Río Calchaquí). En el laboratorio de preparación de microfósiles del Centro de Ecología Aplicada del Litoral (CECOAL), las sedimentitas más finas (pelitas) se sometieron al tratamiento con ácido clorhídrico y ácido fluorhídrico (Volkheimer y Melendi 1976), posteriormente, los residuos fueron montados sobre portaobjetos estándar, utilizando glicerina gelatina y sellados con parafina. Los preparados forman parte de la Colección Paleontológica “Rafael Herbst”, de la Universidad Nacional del Nordeste, en la Subdivisión Preparados Micropaleontológicos, con la sigla CTES-PMP, Localidad Quebrada El Estanque, CTES-PMP N°: 3430, 3424, 3423, 2931, 2930, 2928, 2924, 2910, 2914, 2913, 2912, 2911, 3410, 2909, 3044, 3042, 3041, 3040 (muestras número 1-18). Río Calchaquí, CTES-PMP N°: 2.872 – 2881 (muestras número 1-10). Cabe destacar que se tuvieron en cuenta para el estudio solo aquellos preparados que contenían palinomorfos. Los mismos fueron observados con Microscopio óptico de luz transmitida, modelo Nikon Eclipse-100 con cámara digital incorporada. Se confeccionó el Diagrama Polínico con el programa Tilia (DOS 1.2. Versión ٢.٠.b.٤, Grimm E. ١٩٩١-١٩٩٣) solamente para Quebrada El Estanque debido a que es la localidad que posee mayor diversidad de especies con el objeto de analizar las asociaciones palinológicas con mayor precisión. Para la elaboración del diagrama se ubicaron los niveles sedimentarios más antiguos en la parte inferior. Luego se cargaron las especies de acuerdo a los grandes grupos vegetales: algas, briófitos, pteridofitas, gimnospermas y angiospermas. Con el fin de establecer zonas palinológicas se utilizó el método de agrupamientos (CONNISS) que se basa en el análisis de agrupamientos (Cluster Analysis). Para el análisis de las zonas se tomó en cuenta la diversidad taxonómica (número total de especies en un momento dado) y la frecuencia relativa de sus integrantes. Se adoptó la siguiente escala establecida por Mautino (2010) y se la adaptó a los valores aquí obtenidos, frecuencia menor al 1%: muy escasa; menor al 5%: escasa; menor al 30%: moderada; menor al 60%: abundante; menor al 100%: muy abundante.
RESULTADOS
Las descripciones, determinación sistemática, afinidad botánica, hábito y hábitat de los palinomorfos hallados se encuentran en Horn (2014). En la localidad Río Calchaquí se hallaron 10 especies de palinomorfos que corresponden en un 96,9% a granos de polen de Magnoliophyta y el 3,5% a granos de polen de Gimnospermas (Gnetales) (Tabla 1). Por el hábito y hábitat de las familias actuales afines, se infiere que habrían integrado las paleocomunidades a) bosque higrófilo: estrato arbustivo-arbóreo con presencia de Rhoipites baculatus (Rutaceae) y Polyadopollenites sp. (Fabaceae, Mimosoideae) (Figura 5 L); b) xerófila, se identificaron cuatro estratos: herbáceo Equisetosporites sp. (Ephedraceae) (Figura 5 B), Tubulifloridites antipodica (Asteraceae) (Figura 5 D) y Chenopodipollis chenopodiaceioides (Chenopodiaceae) (Figura 5 K), arbustivo Psilatricolporites sp. (Solanaceae), arbustivo-arbóreo Triorites orbiculatus (Celtidaceae) (Figura 5 H) y arbóreo Striaticolporites gamerroi (Anacardiaceae) (Figura 5 E), Arecipites sp. (Arecaceae) (Figura 5 C); y c) boscosa de altura, con Alnipollenites verus (Betulaceae) (Figura 5 J) como representante.
En la Quebrada El Estanque se hallaron 26 especies de palinomorfos de los cuales el 81,1% corresponde a algas Chlorophyta, el 5,7% a cigósporas de Zygnemataceae, 3,4% a esporas de Polypodiopsida, el 7,5% a granos de Gimnospermas y el 2,6% a granos de Magnoliophyta (Tabla 1). Habrían conformado las paleocomunidades: a) acuática, integrada por Botryococcus braunii (Botryococcaceae) (Figura 4 A), Pediastrum sp. cf. P. boryanum var. longicorne, (Figura 4 B), Pediastrum simplex (Hydrodictyaceae) (Figura 4 C y D), Coelastrum sp. (Coelastraceae) (Figura 4 E), Oedogonium sp. cf. O. cretaceum (Oedogoniaceae) (Figura 4 F), Mougeotia sp. cf. M. laetivirens (Figura 4 G), Spirogyra cf. S. scrobiculata (Figura 4 H), Ovoidites parvus (Figura 4 I), Ovoidites sp. (Figura 4 J) Ovoidites grandis (Figura 4 K), (Zygnemataceae) y Azolla sp. (Azollaceae); b) palustre, estrato herbáceo Muricingulisporis verrucosus (Pteridaceae) (Figura 4 N y M), Polypodiaceiosporites tumulatus (Pteridaceae) (Figura 4 P), Plicatella sp. (Schizaceae) (Figura 4 O), Leptolepidites major (Figura 4 Q); herbáceo epífito Biretisporites crassilabratus (Hymenophyllaceae) (Figura 4 L) y Corsinopollenites exinitenuis (Onagraceae) (Figura 5 I); c) xerófila, estrato herbáceo Tubulifloridites antipodica (Asteraceae), Graminidites sp. (Poaceae) (Figura 5 G); estrato arbustivo Periporopollenites vivianae (Cactaceae); estrato subarbustivo Echiperiporites parviechinatus (Malvaceae) (Figura 5 B) y estrato arbóreo Margocolporites vanwijhei (Fabaceae-Caesalpiniodeae) (Figura 5 F); d) boscosa de altura, con Podocarpidites marwickii (Podocarpaceae) (Figura 5 A).
El análisis del Diagrama polínico de los componentes en los niveles sedimentarios estudiados de la localidad Quebrada El Estanque (Figura 6), permite dividir a la columna estratigráfica en tres zonas polínicas: Zona A (comprende los niveles 18-11), Zona B (comprende los niveles 10-7) y Zona C (comprende los niveles 6-1).
Zona A, se diferencian dos asociaciones: 1) Asociación de algas Chlorophyta: integrada por Botryococcus braunii, Pediastrum sp. cf. P. boryanum var. longicorne y Pediastrum simplex, comprendida en los niveles 18-16 y 14-11. En los tres primeros niveles (18, 17, y 16), las especies mencionadas presentan frecuencias relativas moderadas a abundantes. En cambio, en los niveles 14-11 se observan variaciones. En el caso de Botryococcus braunii, en el nivel 14 su frecuencia es muy abundante (95%) y se encuentra acompañado por Ovoidites parvus y Ovoidites sp. con muy escasas frecuencias relativas (menores al 1%). En el siguiente nivel (13), su frecuencia relativa sigue siendo abundante y además de las especies de Ovoidites, lo acompañan Pediastrum sp. cf. P. boryanum var. longicorne y Pediastrum simplex con frecuencias relativas escasa y moderada respectivamente. En el nivel 12 se encuentran las tres especies que caracterizan la asociación, aunque con frecuencias moderada/abundante, acompañados con muy escasa frecuencia relativa por Mougeotia laetivirens, Reboulisporites fueguensis, Polypodiaceiosporites tumulatus, Muricingulisporis verrucosus, Biretisporites crassilabratus, Corsinipollenites exinitenuis, y Echiperiporites parviechinatus. En el último nivel (11) solamente se encuentra Pediastrum simplex con una frecuencia relativa muy abundante (70%) acompañado de Ovoidites sprigii, Polipodiaceiosporites tumulatus y Periporopollenites vivianae con representación escasa y Podocarpidites marwickii con representación moderada (20%). 2) Asociación de Zygnemataceae y esporomorfos: integrada por Ovoidites sprigii, Ovoidites grandis, Ovoidites sp., Reboulisporites fueguensis, Murcingulisporis verrucosus, Biretisporites crassilabratus y Corsinipollenites exinitenuis con frecuencia relativa moderada, acompañados por Plicatella sp., Polypodiaceiosporites tumulatus y Podocarpidites marwickii con escasa frecuencia relativa.
Zona B, está integrada por la asociación palinológica Zygnemataceae + esporomorfos. Se destaca en esta zona la ausencia de algas Chlorophyta. Comprende los niveles 10-7. En el nivel 10 Ovoidites parvus presenta una frecuencia relativa muy abundante y Ovoidites grandis moderada, están acompañados por Biretisporites crassilabratus y Podocarpidites marwikii con frecuencias escasas. En el nivel 9 Mougeotia laetivirens, Ovoidites parvus, Ovoidites sp., Spirogyra scrobiculata y Corsinipollenites exinitenuis presentan frecuencias relativas moderadas, junto a Biretisporites crassilabratus y Podocarpidites marwikii con escasa representación. En el nivel 8 Mougeotia sp. cf. M. laetivirens, Biretisporites crassilabratus, Polypodiaceiosporites tumulatus, Muricingulisporis verrucosus, Tubulifloridites antipodica y Podocarpidites marwikii presentan frecuencias relativas moderadas, destacándose Podocarpidites marwikii con el 45%. El último nivel (7) se caracteriza por la frecuencia relativa muy abundante de Podocarpidites marwikii (90%) acompañado por escasos representantes de Oedogonium cretaceum, Muricingulisporis verrucosus y Biretisporites crassilabratus.
Zona C, se distinguen tres asociaciones: 1) Asociación de algas Chlorophyta: se observa en los niveles 6-4 y 1. En el nivel 6 se encuentran Botryococcus braunii con frecuencia relativa abundante y las dos especies de Pediastrum ya mencionadas con frecuencias relativas moderadas. En los niveles 5 y 4 a las especies anteriores se suma Coelastrum sp. con frecuencias relativas moderadas. Se destaca en el nivel 4 la frecuencia relativa abundante de Pediastrum simplex y la escasa representación de Botryococcus braunii y Pediastrum sp. cf. P. boryanum var. longicorne. En el último nivel (1) Botryococcus braunii presenta una frecuencia relativa muy abundante (70%), mientras que las especies de Pediastrum tienen frecuencias relativas moderadas. 2) Asociación de Zygnemataceae y esporomorfos: se observa en el nivel 2, con una moderada representación de Ovoidites parvus, O. sprigii, Muricingulisporis verrucosus, Biretisporites crassilabratus, Podocarpidites marwickii, Graminidites sp. y Margocolporites vanwijhei y escasa de Oedogonium cretaceum, Polypodiaceiosporites tumulatus y Tubulifloridites antipodica. 3) Asociación mixta.
Tabla 1. Especies presentes en cada localidad.
|
Taxón |
Afinidad botánica |
Quebrada |
Río Calchaquí |
|
Chlorophyta |
|||
|
Botryococcus braunii Kützing 1849 |
Botryococcaceae |
|
|
|
Coelastrum sp. |
Coelastraceae |
|
|
|
Pediastrum simplex Meyen 1829 |
Hydrodictyaceae |
|
|
|
Pediastrum sp. cf. P. boryanum var. longicorne Reinsch 1867 |
Hydrodictyaceae |
|
|
|
Oedogonium sp. cf. O. cretaceum Zippi 1998 |
Oedogoniaceae |
|
|
|
Zygnemataceae |
|||
|
Mougeotia sp. cf. M. laetivirens (A. Braun) Wittrock 1877 |
Zygnemataceae |
|
|
|
Spirogyra sp. cf. S. scrobiculata (Stockmayer) Czurda 1932 |
Zygnemataceae |
|
|
|
Ovoidites grandis Zippi 1998 |
Zygnemataceae |
|
|
|
Ovoidites parvus (Cookson y Dettman) Nakoman 1966 |
Zygnemataceae |
|
|
|
Ovoidites sprigii (Cookson y Dettmann) Zippi 1998 |
Zygnemataceae |
|
|
|
Ovoidites sp. |
Zygnemataceae |
|
|
|
Bryophyta sensu lato |
|||
|
Reboulisporites fueguensis Zamaloa y Romero 1990 |
|
||
|
Polypodiopsida |
|||
|
Azolla sp. |
Azollaceae |
|
|
|
Biretisporites crassilabratus Archangelsky 1972 |
Hymenophyllaceae |
|
|
|
Muricingulisporis verrucosus Mautino y Anzótegui 1998 |
Pteridaceae |
|
|
|
Plicatella sp. |
Schizaceae |
|
|
|
Polypodiaceiosporites tumulatus Partridge 1973 |
Pteridaceae |
|
|
|
Gimnospermas |
|||
|
Equisetosporites sp. |
|
||
|
Podocarpidites marwickii Couper 1953 |
Podocarpaceae |
|
|
|
Angiospermas |
|||
|
Arecipites sp. |
Arecaceae |
|
|
|
Alnipollenites verus Potonié ex Potonié 1931 |
Betulaceae |
|
|
|
Chenopodipollis chenopodiaceioides (Martin) Truswell 1985 |
Chenopodiaceae |
|
|
|
Corsinipollenites exinitenuis Anzótegui y Cuadrado 1996 |
Onagraceae |
|
|
|
Echiperiporites parviechinatus Anzótegui y Cuadrado 1996 |
Malvaceae |
|
|
|
Graminidites sp. |
Poaceae |
|
|
|
Margocolporites vanwijhei Gemerad, Hopping y Muller 1968 |
Fabaceae Caesalpiniodeae |
|
|
|
Periporopollenites vivianae Mautino 2011 |
Cactaceae |
|
|
|
Polyadopollenites sp. |
Fabaceae Mimosoideae |
|
|
|
Psilatricolporites sp. |
Solanaceae |
|
|
|
Rhoipites baculatus Archangelsky 1973 |
Rutaceae |
|
|
|
Striaticolporites gamerroi Archangelsky 1973 |
Anacardiaceae |
|
|
|
Triorites orbiculatus McIntyre 1965 |
Celtidaceae |
|
|
|
Tubulifloridites antipodica Cookson 1947 |
Asteraceae |
|
|

Figura. 4. Localidad Quebrada El Estanque A. Botryococcus sp. cf. B. braunii Kützing 1849., CTES-PMP 2928c: 46,1/96,8. B. Pediastrum sp. cf. P. boryanum (Turpin) Meneghini 1840 CTES-PMP 2928b: 36,9/87. C-D. Pediastrum simplex Meyen 1829, CTES-PMP 2928b: 36,9/87. E. Coelastrum sp. CTESPMP 2931b: 43,5/86. F. Oedogonium sp. cf. O. cretaceum Zippi 1998, CTES-PMP 2924 c: 37,3/92,6. G. Mougeotia sp. cf. M. laetivirens (A. Braun) Wittrock 1877, CTES-PMP 2914c: 41,5/90,4. H. Spirogyra cf. scrobiculata (Stockmayer) Czurda 1932, CTES-PMP 2914a: 43/89,6. I. Ovoidites parvus (Cookson y Dettman) Nakoman 1966, CTES-PMP 2913c: 32/91,9. J. Ovoidites sp. CTES-PMP 2914a: 39,8/88,1. K. Ovoidites grandis Zippi 1998 CTES-PMP 2913a: 38,9/95,2. L. Reboulisporites fueguensis Zamaloa y Romero 1990 CTES-PMP 2911b: 34,1/99,3. M. Muricingulisporis verrucosus Mautino 1998, CTES-PMP 2910d: 37/88,5. N. Muricingulisporis verrucosus Mautino 1998, vista ecuatorial, CTES-PMP 2910d: 41,1/89,5. O. Plicatella sp. CTES-PMP 3044a: 33,4/89. P. Polypodiaceiosporites tumulatus Partridge 1973, CTES-PMP 2910c: 40,9/88,2. Q. Leptolepidites major Couper 1958, CTES-PMP 2911b: 30,9/97,4. Escala gráfica: A = 9 µm, B = 5 µm, C, D y H = 7 µm, E= 7 µm, F = 20 µm, G y O = 26 µm, I y P = 13 µm, J y M = 16 µm, k = 28 µm, L = 25 µm, N = 15 µm.
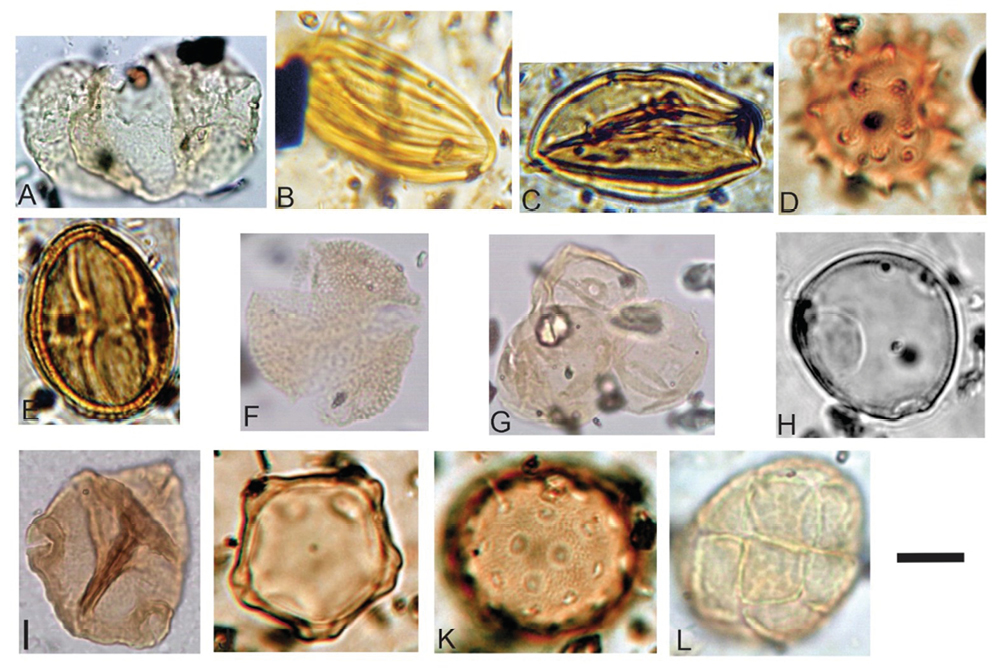
Figura 5. A. Localidad Quebrada El Estanque. Podocarpidites marwickii Couper 1953, CTES-PMP 2924a: 46,5/90,3. B-E. Localidad Río Calchaquí. B. Equisetosporites sp. CTES-PMP 2875a: 34,2/107. C. Arecipites sp. CTES-PMP 2872b: 34,3/103. D. Tubulifloridites antipodica Cookson ex Potonié CTES-PMP 2874a: 37,7/102,1. E. Striaticolporites gamerroi Archangelsky 1973 CTES-PMP 2875b: 33,3/101,1. F-G. Localidad Quebrada El Estanque. F. Margocolporites vanwijhei Germeraad, Hopping y Muller 1968 CTES-PMP 3424a: 38,2/88,8. G. Graminidites sp. CTESPMP 3424a: 41/90,7. H. Localidad Río Calchaquí. Triorites orbiculatus McIntyre 1965 CTES-PMP 2876b: 43,6/106,7. I. Localidad Quebrada El Estanque. Corsinipollenites exinitenuis Anzótegui y Cuadrado 1996 CTES-PMP 2914c: 37,3/90,2. J-L. Localidad Río Calchaquí. J. Alnipollenites verus Potonié ex Potonié 1931 CTES-PMP 2876a: 42,5/103,4. K Chenopodipollis chenopodiaceioides (Martin) Truswell 1985 CTES-PMP 2876a: 36,1/98,1. L. Polyadopollenites sp. CTES-PMP 2875a: 35,5/102. Escala gráfica: A = 12 µm, B, C y D = 5 µm, E = 8 µm, F = 13 µm, G = 0.6 µm, H y J = 7 µm, I = 14 µm, K = 4 µm, L = 11 µm.
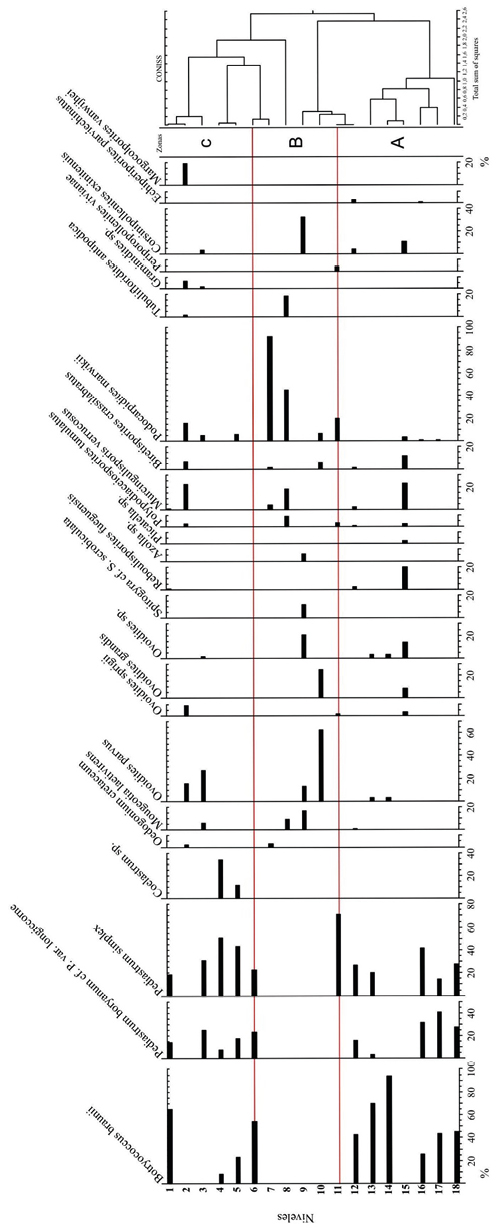
Figura 6. Diagrama polínico de la localidad Quebrada El Estanque (Tilia DOS 1.2. Versión 2.0.b.4, Grimm E. 1991-1993).
discusión
El ambiente de sedimentación de la Formación Palo Pintado es interpretado como un sistema de ríos con sinuosidad intermedia a alta que portaban sedimentos en suspensión y tracción pudiendo desarrollar en algunos sectores un canal y en otros canales múltiples. Este sistema fluvial habría desarrollado amplias llanuras inundables en las que podrían haberse formado lagunas a partir de crecidas o desbordes de los ríos por lluvias excepcionales (Galli et al., 2011; Robledo et al., 2020). Un subambiente palustre habría tenido lugar en la orilla de estas lagunas principalmente en aquellas en proceso de colmatación mientras que en la zona litoral se habría localizado la vegetación de la paleocomunidad palustre herbáceae, mientras que las especies xerófilas se habrían encontrado en zonas más alejadas (Galli et al., 2011). De acuerdo con esta descripción y el análisis de las las características litofaciales, los depósitos sedimentarios en Río Calchaquí corresponden al desarrollo de barras transversales, gravosas y sand wave de depósitos de canal o intracanal, con escaso desarrollo de llanuras inundables (Figura 3). Por lo tanto, la vegetación higrófila se habría desarrollado en las cercanías o la vera de los ríos y la vegetación xerófila en ambientes abiertos del tipo sabana o praderas. En tanto que en Quebrada El Estanque las litofacies corresponden lateralmente al desarrollo de crevasse splay, acreción lateral, lagunas y pantanos en la llanura de inundación (Figura 3). En las orillas de las lagunas habría proliferado la vegetación palustre y en la zona litoral se habría localizado la paleocomunidad palustre herbácea, mientras que las especies xerófilas se habrían encontrado en zonas más alejadas.
Sobre las paleocomunidades inferidas para las dos localidades se destacan las siguientes observaciones: a) En Río Calchaquí no se registran integrantes de cuerpos ácueos y los granos de polen hallados corresponden a vegetación de bosque higrófilo con estrato arbustivo-arbóreo Rhoipites baculatus (Rutaceae) y Polyadopollenites sp. (Fabaceae, Mimosoideae). b) En Quebrada El Estanque el registro palinológico de la paleocomunidad palustre se encuentra relativamente restringido y no se hallaron representantes de bosques de ribera, sin embargo, Anzótegui y Horn (2011) describen impresiones foliares en esta localidad integrantes de dicha paleocomunidad. c) La existencia de una comunidad xerófila en ambas localidades queda confirmada por la presencia de las especies Equisetosporites sp. (Ephedraceae), Tubulifloridites antipodica (Asteraceae), Chenopodipollis chenopodiaceioides (Chenopodiaceae), Psilatricolporites sp. (Solanaceae), Triorites orbiculatus (Celtidaceae), Striaticolporites gamerroi (Anacardiaceae), Arecipites sp. (Arecaceae), Alnipollenites verus (Betulaceae), Graminidites sp. (Poaceae), Periporopollenites vivianae (Cactaceae); Echiperiporites parviechinatus (Malvaceae) y Margocolporites vanwijhei (Fabaceae-Caesalpiniodeae). Hay predominio del estrato herbáceo, siendo Tubulifloridites antipodica (Asteraceae) la única especie que comparten. El registro de Chenopodipollis chenopodiaceioides indicaría la existencia de un ambiente con altas concentraciones de sales (halófitos). d) La paleocomunidad boscosa de altura en ambas localidades se encuentra representada por diferentes especies, Alnipollenites verus (Betulaceae) en Río Calchaquí y Podocarpidites marwickii (Podocarpaceae) en Quebrada El Estanque. Los representantes actuales afines son considerados árboles nativos del noroeste argentino (Zuluaga et al., 2019) lo que justificaría su hallazgo.
Teniendo en cuenta que la mayoría de los taxones actuales afines a las especies que conforman las paleocomunidades de ambas localidades son de origen subtropical, las mismas se habrían desarrollado bajo un clima cálido húmedo con estacionalidad seca.
En relación con el diagrama polínico de la Quebrada El Estanque (Figura 6), se observa la alternancia cíclica de las Asociaciones de a) algas Chlorophyta y b) Zygnemataceae + esporomorfos.
a) La asociación de algas Chlorophyta presenta variaciones en sus frecuencias relativas en los niveles donde se encuentran. Estas variaciones podrían ser interpretadas como indicios de cambios en la concentración de nutrientes del agua. Por ejemplo, la abundancia de Pediastrum simplex en algunos niveles podría deberse a la mayor eutrofización del agua, en cambio la de Botryococcus braunii representaría cuerpos ácueos meso a oligotróficos (Martínez et al., 2008). En términos generales, los tres taxones habrían coexistido en un ambiente de aguas eutróficas a oligotróficas, estables, claras y oxigenadas bajo clima cálido. Botryococcus braunii presenta colonias compuestas sin ramificar y otras ramificadas de color amarillo, anaranjado y marrón. En algunas colonias muy bien preservadas se observan las copas (no así las autoesporas) y en otras los anillos de crecimiento. La mayoría posee buena preservación, aunque en algunos casos las colonias se presentan como masas amorfas. Estas características reflejan diferentes estadios de desarrollo (jovenes y/o adultas) que se habrían producido bajo condiciones ambientales y/o climáticas variables durante un largo período de tiempo y sufriendo un rápido enterramiento en ausencia de oxígeno (Guy Olshon, 1992; Rodríguez Amenábar y Ottone, 2003). Cabe destacar que la abundante representación de Pediastrum simplex, acompañado de Coelastrum sp. indicarían aguas eutróficas a mesotróficas más cálidas (Jankovská y Komárek, 2000).
b) Asociación de Zygnemataceae + esporomorfos. La presencia de cigósporas de Zygnemataceae (en ausencia de algas Chlorophyta) indica un ambiente de estrés hídrico, ya sea por disminución de la concentración de oxígeno en el agua o por desecación (Van Geel y van der Hammen, 1978). En el ambiente terrestre la diversidad de especies de Polypodiopsida se interpreta como eventos de colonización o avance de especies oportunistas (Llorens, 2005). Corsinipollenites exinitenuis (Onagraceae) habría acompañado a estos helechos conformando la paleocomunidad palustre. El registro abundante de Podocarpidites marwikii en esta asociación obedecería a la capacidad que muestra la especie actual afín (Podocarpus parlatorei) de colonizar sitios abiertos de altura y disturbados (Blendinger, 2006; Llorens, 2005; Mautino, 2010). Integran esta asociación en forma esporádica taxones xerófilos de ambientes abiertos tales como, Tubulifloridites antipodica (Asteraceae), Graminidites sp. (Poaceae), Echiperiporites densiechinatus (Malvaceae) y Margocolporites vanwijhei (Fabaceae).
La región de los valles Calchaquíes durante el depósito de los sedimentos de la Formación Palo Pintado (en el intervalo Mioceno Medio-Plioceno) fue afectada por la elevación andina, los cambios en la circulación oceánica y atmosférica y el volcanismo. Estos factores contribuyeron al establecimiento de la vegetación xerófila desde el Mioceno Medio. El ascenso andino, actuó como barrera, impidiendo el paso de los vientos húmedos procedentes del Atlántico condensado la humedad en el flanco oriental de los Andes propiciando el desarrollo local de bosques higrófilos en el intervalo Mioceno Medio-Plioceno (Barreda et al., 2007; Starck y Anzótegui, 2001). De acuerdo con el mapa paleofitogeográfico de Argentina para el lapso Mioceno Tardío-Plioceno de Barreda et al., (2007) esta palinoflora integra la Provincia Paleofitogeográfica Neotropical Neógena, definida a partir de la presencia de taxones Neotropicales y donde la vegetación sería semejante a la presente en la actual Provincia Chaqueña. Se reconocen así elementos que en la actualidad se encuentran en las Provincias Fitogeográficas del centro norte de la Argentina (de las Yungas, Paranaense, Chaqueña, del Monte y del Espinal), por lo que podría considerarse a esta palinoflora como los antecesores de las mismas.
conclusiones
La palinoflora hallada está compuesta por 36 taxones fósiles que pertenecen a colonias de algas Chlorophyta, cigósporas de Zygnemataceae, esporas de Bryophyta y Polypodiopsida, granos de polen de Gimnospermas y Angiospermas. Considerando el hábito y hábitat de las especies actuales afines a los taxones fósiles se han identificado en Río Calchaquí las paleocomunidades bosque higrófilo, xerófila y bosques de altura y en Quebrada El Estanque las paleocomunidades acuática, palustre, xerófila y bosque de altura.
De acuerdo a la asociación de litofacies y elementos arquitecturales los sedimentos de Río Calchaquí corresponden al desarrollo de barras transversales, gravosas y sand wave de depósitos de canal o intracanal, con escaso desarrollo de llanuras inundables. En tanto que en Quebrada El Estanque las litofacies corresponden al desarrollo lateral de crevasse splay, acreción lateral, lagunas y pantanos en la llanura de inundación. En Río Clachaquí la vegetación higrófila se habría desarrollado en las cercanías o la vera de los ríos y la vegetación xerófila en ambientes abiertos del tipo sabana o praderas. En tanto que en Quebrada El Estanque en las orillas de las lagunas habría proliferado la vegetación palustre y en la zona litoral se habría localizado la paleocomunidad palustre herbácea, mientras que las especies xerófilas se habrían encontrado en zonas más alejadas. En ambas localidades en las áreas más altas se habría prosperado el bosque de altura.
En Quebrada El Estanque, a lo largo de la columna estratigráfica, alternan cíclicamente cuerpos ácueos con condiciones meso- a eutróficas y buena disposición de oxígeno (indicado por la presencia de algas Chlorophyta) con otros que denotan condiciones de estrés hídrico. Los primeros fueron numerosos y prolongados (por la presencia de colonias ramificadas y con distintos estadios de desarrollo de Botryococcus braunii). En aquellos niveles que habrían sufrido estrés hídrico (con cigósporas de Zygnemataceae) se hallaron también esporomorfos de la vegetación circundante que habrían conformado las paleocomunidades terrestres.
En ambas localidades la vegetación se habría desarrollado bajo un clima cálido con estacionalidad seca, esto lo confirma la presencia de la comunidad xerófila con especies como por ejemplo Equisetosporites sp. (Ephedraceae), Tubulifloridites antipodica (Asteraceae), Chenopodipollis chenopodiaceioides (Chenopodiaceae). Esta palinoflora concuerda con la Provincia Paleofitogeográfica Neotropical Neógena (Barreda et al., 2007). Desde el punto de vista taxonómico Spirogyra scrobiculata (Zygnemataceae) se determina por primera vez para la Formación Palo Pintado.
Conflicto de intereses
Las autoras declaran no tener conflicto de intereses.
Agradecimientos
Agradecemos la oportunidad de participar en este volumen especial, para homenajear a dos impulsores y referentes de la paleobotánica argentina, siendo reconocidos también en otros países por sus aportes y dedicación. No solo mantuvimos con ellos una relación laboral, sino también compartimos momentos gratos y especiales que siempre recordamos con mucho cariño.
A la Dra. Luisa Anzótegui por la oportunidad que me brindó en los viajes de campo de conocer un ser absolutamente excepcional y compartir trabajos y discusiones muy enriquecedoras (CG).
Agradecemos a los revisores por sus observaciones que contribuyeron a mejorar sustancialmente este trabajo. Por las mismas razones agradecemos también al Dr. Juan Manuel Robledo.
Contribuciones de los autores
La Dra. M.Y. Horn contribuyó con el estudio palinológico de las dos localidades fosilíferas e información desarrollada en su tesis doctoral. Por su parte la Dra. C.I. Galli aportó el análisis de las litofacies y estructuras asociadas, el perfil estratigráfico de los afloramientos de la Formación Palo Pintado y las figuras 1, 2, y 3.
Acevedo, T.L., Mautino, R.L., Anzótegui, L.M. y Cuadrado G.A. (1997). Estudio palinológico de la Formación Palo Pintado (Mioceno superior), Provincia de Salta, Argentina. Parte II: Esporas. Geociências, 2, 112-120.
Anzótegui, L.M. (1998). Hojas de angiospermas de la Formación Palo Pintado, Mioceno superior, Salta, Argentina. Parte 1: Anacardiaceae, Lauraceae y Moraceae. Ameghiniana, 35, 25-32.
Anzótegui, L.M. (2006) Paleofloras del Mioceno en los Valles Calchaquíes, Noroeste de Argentina [Tesis Doctoral, Universidad Nacional del Nordeste].
Anzótegui, L.M. y Cuadrado, G.A. (1996). Palinología de la Formación Palo Pintado, Mioceno superior, Provincia de Salta, República Argentina. Parte I: Taxones nuevos. Revista Española de Micropaleontología, 28, 77-92.
Anzótegui, L.M. y Horn, M.Y. (2011). Megaflora de la Formación Palo Pintado (Mioceno Superior) Salta, Argentina. Parte II. Revista Brasileira de Paleontología, 14(3), 239-254.
Anzótegui, M.L., Mautino, L.R., Garralla, S.S., Herbst, Robledo, J.M. y Horn, M.Y. (2017). Paleovegetación cenozoica del noroeste argentino. En: Ciencias de la Tierra y Recursos Naturales del NOA. Relatorio del XX Congreso Geológico Argentino, 767-781.
Anzótegui, M.L, Mautino, L. R., Horn, M.Y., Garralla, S.S., Robledo, J.M. (2019). Paleovegetación del Mioceno tardío del noroeste de Argentina. Opera Lilloana, 109-130.
Armella M.A., Esteban G.I., García-López D.A. y Nasif N.L. (2016). Vertebrados de la Formación Palo Pintado (Mioceno superior-Plioceno inferior) en la localidad de Angastaco, Provincia de Salta, Argentina [XXX Jornadas Argentinas de Paleontología de Vertebrados]. Actas 156.
Barreda, V., Anzótegui, L.M., Prieto, A.R., Aceñolaza, P., Bianchi, M.M., Borromei, A.M., Brea, M., Caccavari, M., Cuadrado, G.A., Garralla, S.S., Grill, S.G., Guerstein, R., Lutz, A.I., Mancini, M.V., Mautino, L.R., tone, E.G., Quattrocchio, M., Romero, E.J., Zamaloa, M.C. y Zucol, A. (2007). Diversificación y cambios de las Angiospermas durante el Neógeno en Argentina. Asociación Paleontológica Argentina, Publicación Especial 50° Aniversario, Ameghiniana 11: 173-191.
Bona P., Starck D., Galli, C.I., Gasparini, Z. y Reguero, M. (2014). Caiman cf. Latirostris (Alligatoridae, Caimaninae) in the late Miocene Palo Pintado Formation, Salta province, Argentina: Paleogeographic and Paleoenvironmental considerations. Ameghiniana 51(1), 26-36.
Blendinger, P.G. (2006). El pino del cerro (Podocarpus parlatorei) como modelo de estudio para la conservación, ecología y biogeografía de los bosques montanos nublados de las yungas. LIEY Ecología Subtropical, Laboratorio de investigaciones ecológicas de las Yungas, Facultad de Ciencias Naturales e Instituto Miguel Lillo Universidad Nacional de Tucumán.
Candela A. M., Galli, C.I. Rasia L.L y Voglino D. (2021). Late Miocene mammals from the Calchaquí Valley (Palo Pintado Formation, northwestern Argentina): Biogeographic and paleoenvironmental implications for the southern central Andes. Journal of South American Earth Sciences 112:103602 https://doi.org/10.1016/j.jsames.2021.103602
Díaz, J.I. (1985). Análisis estratigráfico del Grupo Payogastilla, Terciario Superior del Valle Calchaquí, provincia de Salta, República Argentina [4° Congreso geológico chileno]. Actas 211-234.
Díaz, J.I. (1987). Estratigrafía y sedimentología del Terciario Superior de la región comprendida entre los valles de los ríos Calchaquí y Guachipas, Provincia de Salta. [Tesis doctoral, Universidad Nacional de Tucumán].
Díaz, J.I. y Malizzia, D.C. (1983). Estudio Geológico y sedimentológico del Terciario Superior del Valle Calchaquí (Departamento de San Carlos, Salta). Boletín Sedimentológico, 2(1), 8-21.
Díaz, J.I., Malizzia, C.D. y Bossi, G. (1987). Análisis estratigráfico del Grupo Payogastilla [10° Congreso Geológico Argentina]. Actas II, 113-116.
Galli, C.I., Vides, M.E. y Flores, P. (2010). Sedimentological and climatic control in the clay minerals distribution in the fluvial deposits of the Palo Pintado Formation (upper Miocene), Salta Province, Argentina [18° International Sedimentological Congress Sedimentology at the Foot of the Andes]. Actas: 376.
Galli, C.I., Anzótegui, L.M., Horn, M.Y. y Morton, L.S. (2011). Paleoambiente y paleocomunidades de la Formación Palo Pintado (Mioceno-Plioceno), Provincia de Salta, Argentina. Revista Mexicana de Ciencias Geológicas, 28(1), 161-174.
Galli, C.I. y Reynolds, J. (2012). Evolución paleoambiental del Grupo Payogastilla (Eoceno - Plioceno) en el valle Calchaquí - Tonco, provincia de Salta, Argentina. Relatorio de la XIII Reunión Argentina de Sedimentología, Salta: 67-80.
Galli C.I., R.N. Alonso, B.L. y Coira, Herrera Oviedo E.P., (2017). Las cuencas de antepaís Cenozoicas de Cordillera Oriental, Noroeste Argentino. Ciencias de la Tierra y Recursos Naturales del NOA, Relatorio del XX Congreso Geológico Argentino - Tucumán 2017, pp. 209-236.
Galli C.I., B. Coira, R. Alonso, J. Reynolds, M. Matteini, N. Hauser, (2014 a). Tectonic controls on the evolution of the Andean Cenozoic foreland basin: evidence from fluvial system variations in the Payogastilla Group, in the Calchaquí, Tonco, and Amblayo Valleys, NW Argentina. Journal of South American Earth Sciences, 52: 234–259.
Galli C. I., B. Coira, R. N. Alonso, M. Matteini, N. Hauser, (2014 b). El Grupo Payogastilla (Cenozoico) en los valles Calchaquí, Tonco y Amblayo, provincia de Salta, Argentina. Acta Geológica Lilloana, 26 (1): 30–52.
Garralla S.S., Anzótegui, L.M. y Mautino, L.R. (2016). Relaciones paleoflorísticas del Mioceno-Plioceno del norte argentino. PeAPA, 16(1), 1-13.
Grimm, E. (1991). Tilia Software. Illinois State Museum. Research and Collection Center Sprigfield Illinois.
Guy-Ohlson, D. (1992). Botryococcus as an aid in the interpretation of paleoenviroment and depositional processes. Review of Paleobotany and Palynology, 71, 1-15.
Herbst, R., Anzótegui, L.M. y Jalfin, G. (1987). Estratigrafía, paleoambientes y dos especies de Salvinia Adanson (Filicopsida) del Mioceno superior de Salta, Argentina. Revista FACENA, 7, 15-42.
Herbst, R., Anzótegui, L.M., Esteban, G., Mautino, L.R., Morton, L.S. y Nassif, N. (2000). Síntesis paleontológica del Mioceno de los valles Calchaquíes, noroeste argentino. En: Aceñolaza, F. y Herbst, R. (Eds.), El Neógeno de Argentina. Serie Correlación Geológica: 263-288.
Hongn, F., Seggiaro, R., 2001, Hoja Geológica 2566-III, Cachi: Provincias de Salta y Catamarca. Instituto de Geología y recursos Minerales, Servicio Geológico Minero Argentino. Buenos Aires, Boletín No. 248, 87 pp.
Horn, M.Y. (2014). Palinofloras de las formaciones El Morterito y Palo Pintado (Mioceno Superior-Plioceno Inferior), noroeste de Argentina [Tesis Doctoral, Facultad de Ciencias Naturales y Museo de la Universidad Nacional de La Plata].
Horn, M.Y., Adami Rodrigues, K. y Anzótegui, L.M. (2011a). Primeras evidencias de interacción insecto-planta en las Formaciones San José y Palo Pintado (Mioceno Medio a Superior), Noroeste de Argentina. Revista Brasileira de Paleontología, 14(1),87-92.
Horn, M.Y., Galli, C.I., Mautino, L.R. y Anzótegui, L.M. (2011 b). Palinología y litofacies de la Formación Palo Pintado (Mioceno Superior), en las localidades Río Calchaquí y Quebrada El Estanque, Salta Argentina. Ameghiniana, Suplemento Resúmenes, 48(4), 15-16.
Jankovská, V. y Komárek, J. (2000). Indicative value of Pediastrum and other Coccal green algae in paleoecology. Folia Geobotanica, 35, 59-82.
Llorens, M. (2005). Palinología de la Formación Punta del Barco, Cretácico Inferior de Santa Cruz [Tesis Doctoral en Ciencias Naturales, Universidad Nacional de La Plata].
Martínez, L. C. A y Lutz, A. I. (2007). Nuevo género y especie de liana del Mioceno Superior (Formación Palo Pintado), provincia de Salta, Argentina. Ameghiniana, 44, 205-213.
Martínez, M.A., Ferrer, N. y Asencio, M. (2008). Primer registro de algas dulceacuícolas del Paleógeno de la Cuenca de Ñirihuau, Argentina: descripciones sistemáticas y análisis palinofacial. Ameghiniana, 45(4), 719-735.
Mautino, L.R. (2010). Palinofloras de las Formaciones San José y Chiquimil (Mioceno Medio y Superior), Noroeste de Argentina [Tesis Doctoral en Biología. Universidad Nacional del Nordeste].
Morton, L.S. (1992). Bivalvos dulceacuícolas de la Formación Palo Pintado (Mioceno Tardío) del valle Calchaquí. FACENA, 9: 77-91.
Morton, L.S. y Herbst, R. (2012). Moluscos fósiles continentals (Bivalvia-Gastrópoda) del NOA Y NEA de Argentina [1º Simposio del Mioceno-Pleistoceno del Centro y Norte de Argentina]. Libro de Resúmenes 26.
Robledo, J.M., Sarzetti, L.C. y Anzótegui, L.M. (2015). Phytophagy on fossil ferns from Argentina (Palo Pintado Formation, late Miocene): a review of their fossil record and ichnotaxonomy. Revista Brasileira de Paleontologia 18(2), 225-238.
Robledo, J.M, Horn, M.Y., Galli, C.I. y Anzótegui, L.M. (2020). Inferencias paleoclimáticas para el Mioceno Tardío de la Cuenca de Angastaco basadas en el análisis fisionómico foliar: Formación Palo Pintado, Salta, Argentina. Andean Geology, 47(2), 418-429. doi: 10.5027/andgeoV47n2-3231.
Robledo, J.M, Barasoain Goñi, D, Quiñones, S.I, Landa Ramirez, E.M, Zurita, A.E y Galli, C.I. (2023). Reconstructing the late miocene paleoenvironment of northwestern Argentina: new sedimentological and faunistic evidence from the Palo Pintado formation. Rivista Italiana di Paleontologia e Stratigrafia, 129(2), 287-305. doi:10.54103/2039-4942/18781 ISSN: 0035-6883.
Rodríguez Amenábar, C. y Ottone, E.G. (2003). La aplicación de Botryococcus (Chlorococcales) como indicador paleoambiental en el Triásico de Argentina. Revista Española de Micropaleontología, 35(2), 161-169.
Salfity, J.A., Monaldi, C.R., 2006, Hoja Geológica 2566-IV, Metán: Programa Nacional de Cartas Geológicas de la República Argentina, 319, 74 pp.
Starck, D. y Anzotegui, L. (2001). The late Miocene climatic change persistence of a climatic signal through the orogenic stratigraphic record in northwestern of Argentina. Journal South American Earth Sciences 14: 763-774, doi: 10.1016/S0895-9811(01)00066-9.
Van Geel, B. y Van Der Hammen (1978). Zygnemataceae in Quaternary Colombian sediments. Review of Paleobotany and Palynology, 25, 377-392.
Volkheimer, W. y Melendi, D. (1976). Palinomorfos como fósiles guía (3ª. Parte). Técnicas de Laboratorio palinológico. Revista Minera, Geología y Mineralogía, 34, 19-30.
Zimicz N., Payrola P. y del Papa C. (2018). New, Late Miocene mammalian assemblage from the Palo Pintado Formation (Northwestern Argentina). Journal of South American Earth Sciences, 81: 31-44.
Zuloaga, F. O., Belgrano, M. J., y Zanotti, C. A. (2019). Actualización del Catálogo de las Plantas Vasculares del Cono Sur. Darwiniana, Nueva Serie, 7(2), 208–278. https://doi.org/10.14522/darwiniana.2019.72.861. Apéndice 1 http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/861/1167.
Zurita, A.E., Galli C.I., Barasoain, D., Quiñones, S.I., Luna, C.A., Candela, A.M., Landa Ramírez, E.M. and Alonso, R.N. (2024) New Cingulata (Mammalia, Xenarthra) from the late Neogene Palo Pintado Formation (Northwestern Argentina). Journal of South American Earth Science, 150, 105233. https://doi.org/10.1016/j.jsames.