Articulo original
Original article
35 (2) | Jul - Dic, 2025
Open Access: https://revistas.unne.edu.ar/index.php/fce
E-mail: revistafacena@exa.unne.edu.ar
New Lower Cretaceous ferns (Polypodiopsida) from the Aptian Baqueró Group, Santa Cruz Province, Argentina
Nuevos helechos (Polypodiopsida) del Cretácico Inferior del Grupo Baqueró (Aptiano), Provincia de Santa Cruz, Argentina
Vera, Ezequiel Ignacio * ¹ 4, Perez Loinaze, Valeria S. ¹ 4 , Passalia, Mauro Gabriel 2 4
& Llorens, Magdalena 3 4
1. Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, CONICET. Buenos Aires, Argentina.
2. INIBIOMA, CONICET-UNCo. San Carlos de Bariloche, Río Negro, Argentina.
3. Laboratorios de Palinología y de Botánica, Facultad de Ciencias Naturales, Universidad Nacional de la Patagonia. Trelew, Chubut, Argentina.
4. Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina.
* Autor de correspodencia: evera@macn.gov.ar
Recibido/Received: 23 Enero, 2025 | Aceptado/Acepted: 07 de Abril, 2025 | Publicado/Published: 05 de Nov., 2025
Editores invitados: Dra. Silvia Gnaedinger & Dr. Ángel R. Miño-Boilini
Como citar este artículo: Vera, E. I., Perez Loinaze, V. S., Passalia, M. G. & Llorens, M. (2025). New Lower Cretaceous ferns (Polypodiopsida) from the Aptian Baqueró Group, Santa Cruz Province, Argentina. Revista FACENA 35(2), 22-45. Doi: https://doi.org/10.30972/fac.3528826
Resumen: El Grupo Baqueró (Aptiano) contiene una de las floras fósiles mejor documentadas del Cretácico Inferior de Patagonia, la cual consiste de una asociación diversa de impresiones/compresiones de hojas, ramas, conos, madera silicificada y carbonizada, mesofósiles y palinomorfos aislados, representando la mayoría de los grandes clados de plantas. Entre estos, los helechos son el grupo más diverso, y las angiospermas recuperadas muestran algunos de los primeros pasos en la evolución del clado. La flora fósil de esta unidad ha sido objeto de estudios ininterrumpidos desde 1960, y su abundancia permite aún hoy el descubrimiento de novedades paleontológicas. En esta contribución, se reportan tres nuevos taxones de helechos para la unidad inferior del Grupo Baqueró (i.e., Formación Anfiteatro de Ticó) sobre la base de impresiones/compresiones de hojas. Paramatonia rafaherbstiana sp. nov. es un helecho estéril morfológicamente comparable a Matonia y Phlebopteris, y representa el primer posible macrofósil de Matoniaceae para la unidad. Un segundo taxón, identificado como Cyatheaceae indet., es un fragmento de fronde fertil, con pínnulas que portan indusios con forma de copa (cyatheoides). El tercer taxón reconocido es referido como Polypodiopsida indet., y se caracteriza por poseer pínnulas pequeñas dentadas portando hasta 10 soros en su cara abaxial. Estos nuevos taxones incrementan la diversidad de helechos para el Grupo Baqueró.
Palabras claves: Formación Anfiteatro de Ticó; Matoniaceae; Cyatheaceae; Paramatonia; Patagonia.
Abstract: The Aptian Baqueró Group contains one of the most well-known Lower Cretaceous fossil floras of Patagonia, consisting of a rich assemblage of impression/compressions leaves, twigs, cones, wood silicifications and carbonizations, mesofossils and isolated palynomorphs. This assemblage represents most major clades of plants, with ferns being the most diverse group, and the recovered angiosperms show some of the first steps in the evolution of the clade. The fossil flora of this unit has been the focus of continuous studies since 1960, and its abundance still allows for the discovery of paleontological novelties. In this contribution, three new fern taxa are reported from the lower unit of the Baqueró Group (i.e. Anfiteatro de Ticó Formation) based on impression/compressions of leaves. Paramatonia rafaherbstiana sp. nov. is a sterile fern morphologically comparable to Matonia and Phlebopteris, and represents the first putative Matoniaceae macrofossil from the unit. A second taxon, identified as Cyatheaceae indet., is a fertile fragmentary frond with pinnules bearing cup-shaped (cyatheoid) indusia. The third recognized taxon is referred to as Polypodiopsida indet., and is characterized by having small dentate pinnules bearing up to 10 sori on the abaxial side. These new taxa increase the specific fern diversity of the Baqueró Group.
Keywords: Anfiteatro de Ticó Formation; Matoniaceae; Cyatheaceae; Paramatonia; Patagonia.
Introducción
The Baqueró Group outcrops in Patagonia, Argentina, host one of the most diverse and best-known Early Cretaceous floras in the Southern Hemisphere. This Aptian assemblage comprises palynomorphs, mesofossils, leaf impression/compressions and fossil woods of most major plant clades, including non-vascular plants, lycopsids, ferns, equisetaleans, conifers, cycads, bennettites, ginkgoaleans, and primitive angiosperms (see Archangelsky, 2003; Del Fueyo, 2007; Limarino et al., 2012; Villar de Seoane and Archangelsky, 2014, and cites therein). Numerous contributions have been published since 1960 focused on the fossil content of the “Ticó Flora”-bearing unit, originally named as Baqueró Formation, which was later elevated to the rank of Group, and subdivided, from base to top, in the Anfiteatro de Ticó, Bajo Tigre and Punta del Barco formations (Cladera et al., 2002).
The first palynological studies of this flora were published by Archangelsky and Gamerro (1965, 1966a, b, c), on palynofloras recovered from the Anfiteatro de Ticó Formation. Subsequent research on the same unit was conducted by Archangelsky and Villar de Seoane (1992, 1994, 1996, 1998, 2005), Archangelsky and Archangelsky (2013) and Villar de Seoane and Archangelsky (2014). Palynofloras from the Punta del Barco Formation were analyzed by Llorens (2003, 2008a, b, 2012), while more recent contributions by Llorens and Perez Loinaze (2016) and Perez Loinaze and Llorens (2018) provided additional data on palynofloras from the Anfiteatro de Ticó and Punta del Barco formations. Fossil woods were identified in the Baqueró Group in the pioneer works in the area, but were only recently studied (Vera and Césari, 2012, 2015; Vera and Perez Loinaze, 2022). The abundant and exquisitely preserved megafloristic remains from the Baqueró Group are perhaps the most significative fossils of the unit, and were subject of numerous contributions (e.g., Archangelsky, 2003; Del Fueyo and Archangelsky, 2002, 2005; Villar de Seoane, 2005; Del Fueyo et al., 2007; Passalia et al., 2010; Carrizo et al., 2012; Lafuente Díaz et al., 2021 and cites therein). One of the most diverse plant groups recorded in the unit is the ferns, with Hausmannia papilio Feruglio emend. Herbst 1960 and several species of the “Gleichenites” being among the most iconic and abundant in certain baqueroan strata. These ferns have been the focus of multiple studies by Rafael Herbst (1960, 1962, 1966).
The rich paleobotanical record of the Baqueró Group and its stratigraphic provenance within the unit have been summarized in the works of Archangelsky (2003), Del Fueyo et al. (2007), and Limarino et al. (2012). Despite over sixty years of detailed paleobotanical and palynological research on this unit, the outcrops of the Baqueró Group continue to yield new and noteworthy discoveries, as reported by Llorens and Perez Loinaze (2016), Perez Loinaze and Llorens (2018), Llorens et al. (2020), Lafuente Díaz et al. (2021), Vera and Perez Loinaze (2022), among others. The importance of the Baqueró Group is further heightened by the availability of absolute dating, which constrain the age of the entire Baqueró Group to the upper Aptian (Césari et al., 2011, Perez Loinaze et al., 2013, Passalia et al., 2016).
In this contribution we describe new fossil ferns identified from strata of the Anfiteatro de Ticó Formation at two localities where the Baqueró Group crops out (i.e. Anfiteatro de Ticó and Cerro Bayo localities).
GEOLOGICAL SETTING
The Baqueró Group crops out in the south of Patagonia, in the Macizo del Deseado area, Santa Cruz Province, Argentina (Fig. 1). This group consist, from bottom to top, of the Anfiteatro de Ticó, Bajo Tigre and Punta del Barco formations (Cladera et al., 2002). The Baqueró Group was deposited in continental environments, mostly fluvial and lacustrine, and is composed of sandstones, conglomerates, tuffs and mudstones, reaching 200 m in thickness (Limarino et al., 2012). The basement of the basin includes volcanic deposits from the Chon Aike and La Matilde fomations (Bahía Laura Group, Middle-Upper Jurassic) and continental strata from the Bajo Grande Formation (Upper Jurassic–Lower Cretaceous) (Panza et al., 1998). The Baqueró Group strata are unconformably overlain by the lower Miocene marine Monte León Formation and Paleogene and Neogene basalts (Panza et al., 1998). The Anfiteatro de Ticó Formation consist mainly of conglomerates, cross-bedded sandstones, fine-grained sandstones, and thin interbedded tuffs, all deposited in lacustrine and fluvial environments (Cladera et al., 2002; Limarino et al., 2012). Perez Loinaze et al. (2013) obtained an age of 118.23 ± 0.09 Ma from tuffs in the lower levels of Anfiteatro de Ticó Formation. The overlying unit, the Bajo Tigre Formation, is composed of tuffs, muddy tuffs, tuffaceous sandstones, and conglomerates, deposited in a fluvial environment (Cladera et al., 2002). This unit differs from the underlying unit by the absence of lacustrine systems and the presence of abundant ash-fall deposits (Limarino et al., 2012). Radiometric dating on this unit yielded an age of 116.85 ± 0.26 Ma (Passalia et al., 2016). Finally, the Punta del Barco Formation is composed of tuffs, muddy tuffs, tuffaceous sandstones and conglomerates, interpreted as fluvial systems and ash-fall deposits (Cladera et al., 2002; Limarino et al., 2012). Césari et al. (2011) reported a 206Pb/238U age of 114.67 ± 0.18 Ma for this unit. Later, Passalia et al. (2017) reported an assemblage of putative basal eudicots (aff. Ranunculales) from the uppermost levels of the Punta del Barco Formation, using them as a basis to suggest that this unit may extend to the uppermost Aptian/Albian boundary. For a detailed characterization of the geology of the Baqueró Group and its fossil-bearing levels, see Cladera et al. (2002), Limarino et al. (2012) and Passalia et al. (2016).
MATERIALS AND METHODS
The studied remains were collected from outcrops of the Anfiteatro de Ticó Formation at the Anfiteatro de Ticó and Cerro Bayo localities (Fig. 1), during three field trips conducted in 2008, 2010 and 2011. All the specimens analyzed are housed in the Museo Regional Provincial Padre Manuel Jesús Molina, Santa Cruz province, Argentina, under MPMPB catalog numbers. Megafloristic remains were studied using a Nikon SMZ800 binocular microscope and photographed with an attached camera (Nikon Coolpix 995). Attempts to study details of fertile structures using scanning electron microscopy and fluorescence microscopy were almost unsuccessful. Descriptions are based on Miller and Hickey (2008) and Machado et al. (2021, 2023).
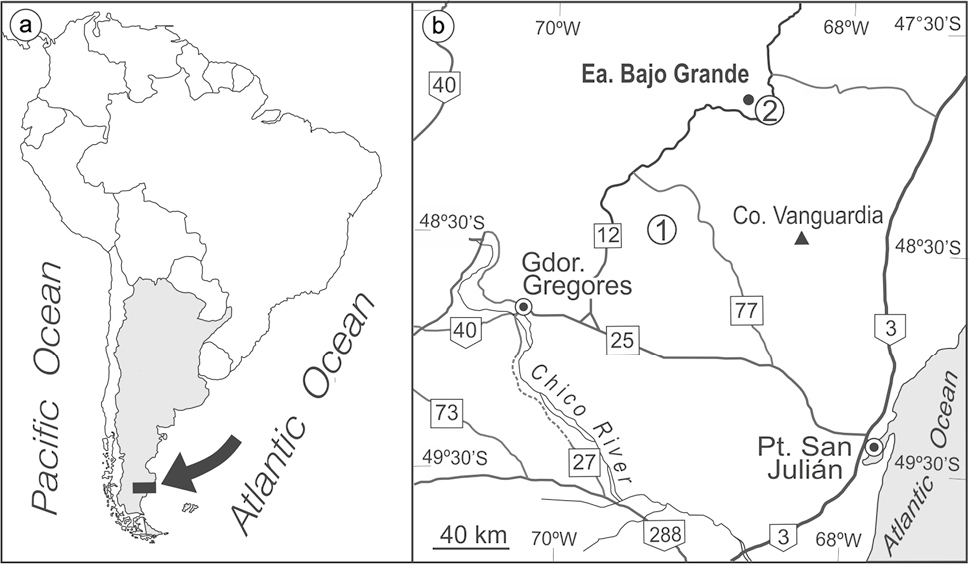
Figure 1. a. Location map of the studied area in the Deseado Massif Basin (black rectangle). b. Detail of the area showing outcrops of Baqueró Group (gray) (based on the geological survey of Panza et al., 1998). 1, Anfiteatro de Ticó locality; 2, Cerro Bayo locality.
results
Systematic Paleontology
Class Polypodiopsida
Order Gleicheniales Schimp. 1869?
Family Matoniaceae C. Presl 1847?
Paramatonia Miller et Hickey 2008
Type species. Paramatonia linearis Miller et Hickey 2008.
Remarks. Fossil ferns with similar morphology have been assigned to the genera Matonidium Schenk, Phlebopteris Brongniart, and even to the extant Matonia R.B. ex Wall. The classification of these genera is based not only on vegetative features, but also on the morphological characterization of their reproductive structures. Miller and Hickey (2008) erected the fossil genus Paramatonia to encompass remains exhibiting features of Matonidium/Phlebopteris/Matonia, for which there was insufficient information to place them definitively within these taxa, but that still have enough morphological features to be recognized as different entities.
Paramatonia rafaherbstiana Vera, Perez Loinaze, Passalia et Llorens sp. nov.
Figure 2.
Synonymy.
2012. Phlebopteris–Matonidium–Matonia complex. Limarino et al., 2012, Fig. 4.9,10.
Etymology. In reference to the late Dr. Rafael Herbst, mentor, colleague, and friend, in recognition of his many contributions to the study of fossil plants from South America, in particular focused on fossil ferns.
Type material. Holotype MPMPB 3192, Paratype MPMPB 3211.
Additional specimens. MPMPB 3193, 3212, 3213, 3219, 3220, 3221.
Type locality. Anfiteatro de Ticó locality, 48°30’35.94” S, 69°14’12.12” W
Type horizon. Anfiteatro de Ticó Formation, Depositional Sequence 1, Stratigraphic Level 5 (sensu Limarino et al., 2012), upper Aptian.
Species diagnosis. Pinnate leaf fragments; rachis straight and stout; pinnules ranging from triangular (l/w ratio c.1) in the basal region, to linear to slightly falcate (l/w ratio c. 5). The margins are entire and apices acute. The pinnule venation consists of a midvein and numerous lateral veins, which are densely arranged and diverge at angles of 60-90°. Lateral veins fork one to four times at different levels, some lateral veins join, forming irregularly distributed areolae. Fertile structures unknown.
Description. The description is based on all available specimens, which are referred to the same taxonomic entity due to their overall similarity in general morphology, pinnule size and shape, and venation pattern. Complete leaves and fertile leaves are unknown. Pinnate leaf fragments, up to 6.2 cm long, 6.4 cm wide (Fig. 2a, e). The rachis is straight and stout, 1.1–2 mm wide, being widest at the basal sections and non-grooved (Fig. 2a, b, e, f). Pinnule attachment ranges from opposite to alternate (Fig. 2a, e). Pinnules variable in shape and size. The basalmost preserved pinnule 2.9 mm long and 2.8 mm wide at its base, with an l/w ratio c.1. It is triangular in shape and attached at angle of 67° (Fig. 2e, f). Pinnule morphology changes abruptly towards the apex of the pinna, becoming linear to slightly falcate (Fig. 2a–c, e–g), gently curving towards the apex. The longest pinnules reach up to 33 mm in lenght, 6.7 mm at their base, with an l/w ratio c. 5. Attachment angles range from 67 to 79°. The margins are entire, abaxially curved (coriaceous?), and the basal region is straight in the basalmost pinnules, becoming acroscopically and basiscopically decurrent towards the apex (Fig. 2a–c, e–g). Pinnule apices are acute (Fig. 2a, b, e, g). Pinnule venation consists of a midvein and numerous lateral veins. The midvein is straight to slightly recurved, ending just below the pinnules apex. Lateral venation with a density of 48–53 veins per cm at the margin (Fig. 2b–d). Lateral veins fork once to four times; with dichotomies occurring immediately after their departure from the midvein or extending across all the length of the lamina. Some lateral veins rejoin with adjacent ones, forming irregularly distributed, discrete areoles (Fig. 2b–d). Lateral veins diverge from the midvein at angles of 64–88° and reach the margin of the pinnule (Fig. 2c-d). The lateral veins are alternate to opposite (Fig. 2b–d). The dissection pattern of the venation remains unknown. Interveinlet interval 0.18–0.26 mm (Fig. 2c-d).
Affinities. The morphology of Paramatonia rafaherbstiana sp. nov. resembles taxa referred to the Matoniaceae, as Phlebopteris and Matonia (Miller and Hickey, 2008, and cites therein). Nevertheless, the absence of preserved reproductive structures makes this referral tentative. Matonisporites, a spore genus related with matoniaceous taxa (Balme, 1995, and cites therein), is recorded in lower proportions in palynological assemblages recovered from the Baqueró Group (Perez Loinaze and Llorens, 2018, and cites therein). Conversely, the genus Cyathidites, present in higher proportions, may have been produced by Matoniaceae as well as by other plant taxa (Balme, 1995; Sajjadi and Playford, 2002; Schrank, 2010; Barreda et al., 2012, among others). This suggests that the presence of matoniaceous ferns in the Baqueró palynological assemblages cannot be disregarded. .
Comparisons. Paramatonia rafaherbstiana sp. nov. differs from the only currently described species of the genus P. linearis Miller et Hickey 2008, from the Winthrop Formation (Albian, USA) in having proportionally shorter pinnules and well-defined lateral veins. P. linearis preserves reproductive structures, which are absent in the Patagonian form (Miller and Hickey, 2008). The macroscopic fossil record of Matoniaceae for the whole South America is sparse (Pons, 1988 and cites therein; Nishida et al., 2014). However, there are some reports of the fertile matoniaceous species Konijnenburgia alata (Halle) Kvaček et Dašková from mid-Cretaceous (Albian) plant communities of southwestern Patagonia, Argentina (see Passalia et al., 2018 and cites therein). With this species, P. rafaherbstiana sp. nov. shares some resemblance in the development of elongated pinnules with a thick midvein. On the other hand, lateral veins are less densely distributed in K. alata. Furthermore, they always divide and anastomose, also presenting higher order veins forming a dense network of areoles, contrasting with what is seen in P. rafaherbstiana sp. nov. Matonia jeffersonii Nagalingum et Cantrill 2006 from the Albian Triton Point Formation of Alexander Island, Antarctica, is a taxon morphologically similar to K. alata, showing a dense network of areoles formed by secondary and higher order veins, as well as preserved sori (Nagalingum and Cantrill, 2006; Passalia et al., 2018), all these features differing from P. rafaherbstiana sp. nov. Noteworthy, the lack of preserved sporangia details or spore content, and the absence of the typically pedate lamina division of Matonia, made Passalia et al. (2018) challenge the placement of M. jeffersonii in the genus Matonia.
To date, no other fern species reported from the Baqueró Group shares morphological similarities with Paramatonia.

Fig. 2. a-g. Paramatonia rafaherbstiana sp. nov. a-d. Holotype MPMPB 3192. a. General view of the specimen. b. Detail of the specimen showing morphology of pinnules. c. Detail showing venation pattern. Notice lateral connections with adjacent veins (arrows). d. Detail of lateral veins. Notice lateral connection with adjacent veins (arrow). e-g. Paratype MPMPB3211. e. General view of the specimen. Notice basal triangular pinnules (arrow). f. Detail of basal pinnules. g. Detail of falcate pinnules. Scale bars: a: 2 cm; b, e: 1 cm; c, f: 3 mm; d: 1 mm; g: 5 mm.
Order Cyatheales A. B. Frank in J. Leunis 1877
Family Cyatheaceae Kaulf 1827
Cyatheaceae indet.
Figure 3a–g.
Studied specimens. MPMPB 3264 A and B (part and counterpart).
Locality. Cerro Bayo locality, Bajo Grande Area, 47° 47’ 31.7” S, 68° 45’ 22.4” W.
Horizon. Anfiteatro de Ticó Formation, middle-upper section of Depositional Sequence 1 (probably corresponding to Stratigraphic Levels 5 or 6) sensu Limarino et al. (2012), upper Aptian.
Description. The description is based on a single fertile specimen, which includes part and counterpart. Complete leaves are unknown. Pinnate to distally pinnatifid leaf fragment (pinnae), imparipinnate, up to 28.8 mm long, 7.5 mm wide, tapering towards the apex (Fig. 3a, b). The rachis is straight and stout, 0.4–0.6 mm wide (Fig. 3a–f), and non-grooved. Pinnules attachment ranges from alternate to subalternate (Fig. 3a, b). Pinnules variable in shape and size. The basalmost preserved pinnule measures 4.1 mm long and 1.8 mm wide at its base, with an l/w ratio c. 2.3; it is oblong and more or less falcate, becoming rounded towards the apex of the pinnae. The attachment angle is 63–69° (Fig. 3a–f). The bases are decurrent, connecting with adjacent pinnules toward the apex of the pinnae. The margins are entire to slightly undulating, coinciding with position of the sori); the apex is rounded. Pinnule venation is observable only in the distalmost pinnules, poorly-defined, consisting of a midvein and lateral veins that fork at least once, apparently without anastomosis (Fig. 3e). Pinnules bear a total of 2–10 abaxial sori, arranged alternately to subopposite on each side of the midvein (Fig. 3a, c, d, f). The indusia are cup-shaped, roughly circular in outline, about 0.6–0.9 mm in diameter, and located abaxially (Fig. 3a, c, d, f). At least eight sporangia per sori, circular, with rings 209–379 µm in diameter (Fig. 3c, d, f, g).
Affinities. Several groups of ferns share the presence of a cup-shaped indusia, as Dicksoniaceae, Thyrsopteridaceae, Cyatheaceae, and some Dennstaedtiaceae (Kramer and Green, 1990; Korall et al., 2007). Nevertheless, in most of these taxa, the sporangia are marginal, and the indusium is formed by both a true indusium and a portion of the lamina, denominated as a false indusium (Korall et al., 2007). Only Cyatheaceae have abaxial, cup-shaped (i.e. cyatheoid) indusia that do not include part of the leaf lamina in its structure (Korall et al., 2007, and cites therein). The sori recognized in Cyatheaceae indet. are abaxial, with the margin of the lamina not involved in the structure of the indusium. These are interpreted as true cyatheoid indusia, suggesting affinities with the scaly tree-ferns. Among this clade of ferns, cyatheoid indusia are recorded in the genera Cyathea and Alsophila (Korall et al., 2007). Due to the lack of sufficient preserved information to allow separating these two genera (e.g. spore ultrastructure), we prefer to leave these materials in open nomenclature.
Comparisons. Putative cyatheaceous ferns are scarce in the Baqueró Group macrofloral assemblage, represented only by Cyathea cyathifolia (Villar de Seoane) Villar de Seoane 1999, a taxon originally referred to the fossil genus Cladophlebis (Villar de Seoane, 1996), but later considered comparable to extant Cyathea. Cyatheaceae indet. differs from Cyathea cyathifolia, reported from the same locality and stratigraphic unit (i.e. Anfiteatro de Ticó Formation), by the presence of pinnules with secondary veins without divisions, and sori without indusia in the latest taxon (Villar de Seoane, 1996, 1999). No other fern from the Baqueró Group preserves cup-shaped indusia (see Archangelsky, 2003, and Del Fueyo et al., 2007).
Order unknown
Polypodiopsida indet.
Figure 3h–m.
Synonymy.
2016. Pteridophyte incertae sedis. Passalia et al., 2016, Fig. 6A.
Studied specimens. MPMPB 3236 A, B, C, D, E, F (6 specimens)
Locality. Cerro Bayo locality, Bajo Grande Area, 47°47’ 31.7” S, 68° 45’ 22.4” W.
Horizon. Anfiteatro de Ticó Formation, middle-upper section of Depositional Sequence 1 (probably corresponding to Stratigraphic Levels 5 or 6) sensu Limarino et al. (2012), upper Aptian.
Description. The description is based on six fragmentary specimens originally coming from a single rock sample, which are referred to the same taxonomic entity due to their general morphological similarity, pinnule size and shape, and venation pattern. Complete leaves are unknown. Pinnate leaf fragments (Fig. 3h, j–l), possibly at least bipinnate (see MPMPB 3236 B, Fig. 3k), up to 16 mm long and 9 mm wide. The rachis is straight to flexuous, 0.3–0.4 mm wide, and non-grooved (Fig. 3h, j, k). Pinnule attachment ranges from alternate to subopposite. Pinnules are 4.5 mm long and 0.9 mm wide, with an l/w ratio c. 5; they are oblong to slightly falcate, attached at angles of 55–70°. The basiscopic margin of the pinnules bases is decurrent, but a constriction may be present immediately below the first pair of basal lobes. The margins are toothed or lobed, abaxially curved (coriaceous?), and five rounded lobes on each side of the pinnule, with the basalmost lobes typically the largest. The pinnule apex is rounded, and slightly oriented apically (Fig. 3h–l). Pinnule venation consists of a midvein and lateral veins. The midvein is straight to slightly recurved, ending below the pinnule apex (Fig. 3i, j, l, m). Lateral venation consists of five veins on each side (coinciding with the teeth), which fork once at approximately the same level as the marginal incisions (Fig. 3j, l, m). Lateral veins diverge from the midvein at angles of 40–65° and reach the margin of the pinnule. Lateral veins alternate; dissection pattern of venation catadromous (Fig. 3h–k). The fertile structures are not clearly preserved, but are located abaxially, with up to 10 per pinnule. Each fertile structure consists of a single circular sorus in each lobe, located near the dichotomy of the lateral veins (Fig. 3i, l, m).
Affinities. While its referral to the Polypodiopsida is granted by the presence of fertile structures abaxial to the pinnules lamina, morphology of the frond, and the lack of detailed information of the structure of sori, sporangia and spores precludes further comparisons.
Comparisons. Several fern taxa have been reported from the Baqueró Group that are worth being compared with Polypodiopsida indet. Species originally referred to “Gleichenites”, including Korallipteris argentinica (Berry) Vera et Passalia 2012, Korallipteris feruglioi (Herbst) Vera et Passalia 2012, Korallipteris vegagrandensis (Herbst) Vera et Passalia 2012 and Gleicheniaceaephyllum san-martini (Halle) Vera et Passalia 2013, exhibit proportionally wider and shorter pinnules with entire margins, and bear up to 4 sporangia (Herbst, 1962). Cyathea cyathifolia has simple pinnules with entire margins and simple, unforked, secondary veins (Villar de Seoane, 1996, 1999), in contrast to the lobate pinnules with forked secondaries observed in Polypodiopsida indet. The presence of pinnules with entire margins and cup-shaped indusia in Cyatheaceae indet. described here allows separating it from Polypodiopsida indet. The species defined by Herbst (1966) as Baqueroites padulae has pinnules with entire margins bearing marginally located fertile structures (Herbst, 1966; Carrizo et al., 2018), which contrast to the abaxial position of these structures in the materials described here. Species of the fossil genus Cladophlebis Brongniart reported from the Baqueró Group, apart from lacking information about the reproductive structures, differ from Polypodiopsida indet. by having larger pinnules, entire to slightly undulating margin, and typically by having secondary venation with more proximal divisions and with some cases exhibiting more than one division (Herbst, 1971; Archangelsky, 2003; Carrizo et al., 2012).

Figure 3. a-g. Cyatheaceae indet. a. General view of specimen MPMPB 3264-A. b. General view of specimen MPMPB 3264-B. c-d. Detail of MPMPB 3264-A, showing larger pinnules bearing abaxial sori with cup-shaped indusia. Arrows indicate sori with preserved sporangia. e. Detail of MPMPB 3264-B, showing distal pinnules with preserved primary and lateral veins. f. Detail of MPMPB 3264-A, showing distal pinnules bearing sori. g. SEM image of a pinnule of specimen MPMPB 3264-A, showing partially preserved sporangia rings (arrow). h-m. Polypodiopsida indet. h. General view of specimen MPMPB 3236-A; i. detail of specimen MPMPB 3236-B, showing morphology of the pinnules and position of sori (arrows). j. General view of specimen MPMPB3236-C, showing lateral veins (arrows). k. General view of specimen MPMPB3236-B, showing aligned portions of pinnae. l. General view of specimen MPMPB 3236-E, showing position of sorus (arrow). m. Detail of specimen MPMPB 3236-E showing forking lateral veins (arrows). Scale bars: a, k: 5 mm; b, h: 3 mm; c-f, i-j, l-m: 1 mm; 1 mm; g: 200 µm.
CONCLUSIONS
ACKNOWLEDGMENTS
This work is dedicated to the memory of Dr. Rafael Herbst, in recognition of his numerous contributions to the study of South America’s fossil ferns, among other fossil groups. Beyond his extensive scientific achievements, we also wish to acknowledge him for his generosity in sharing knowledge and experience with students and young colleagues, always with his characteristic sense of humor and enthusiasm. We also express our gratitude to the late Dr. Sergio Archangelsky, a pioneer in the study of the Baqueró Flora, for sharing his knowledge, information and field notebooks with us, which were critical to carry on our studies on this emblematic flora. We are also greatly indebted to Dr. Silvia Césari, not only for coordinating the group during the study of the Baqueró flora, as well as assisting in field and laboratory work but also by having a key role at academic development. Dr. Oscar Limarino kindly participated in several field trips where samples of the Baqueró flora were collected, and his work was critical for understanding the geologic context of the fossil flora of the Baqueró Group. We also thank the two anonymous reviewers that provided suggestions and corrections that enhanced the final version of the manuscript. We extend our gratitude to the Secretaría de Cultura de la Provincia de Santa Cruz for permitting us fieldwork in the area. Finally, we wish to acknowledge all members of the public scientific systems in Argentina, including CONICET and the various universities, who, despite the current challenges by financial cuts, aggression and questioning from different parts of the government and allied forces, continue to produce with effort scientific works of national and international renown, believing that the scientific and technological advances are critical for the development of the country. This work was carried out under research grant PIP 1814 (Consejo Nacional de Investigaciones Científicas y Técnicas).
CONFLICT OF INTEREST
Authors don’t have any conflict of interest to declare.
AUTHOR CONTRIBUTIONS
Vera: Sample collecting, Conceptualization, Formal analysis, Investigation, Original draft preparation, Visualization; Writing - Review & Editing.
Perez Loinaze: Sample collecting, Conceptualization, Investigation, Original draft preparation, Visualization; Writing - Review & Editing.
Passalia: Sample collecting, Investigation, Resources, Writing - Review & Editing.
Llorens: Sample collecting, Resources, Writing - Review & Editing.
Referencias
Archangelsky, S. (2003). La flora cretácica del Grupo Baqueró, Santa Cruz, Argentina. En Monografías del Museo Argentino de Ciencias Naturales. Buenos Aires, pp. xiv þ CD.
Archangelsky, S. & Archangelsky, A. (2013). Aptian angiosperm pollen from the Ticó Flora, Patagonia, Argentina. International Journal of Plant Sciences, 174, 559–571. https://doi.org/10.1086/669928
Archangelsky, S. & Gamerro, J. C. (1965). Estudio palinológico de la Formación Baqueró (Cretácico), provincia de Santa Cruz. Ameghiniana, 4(5), 159–170.
Archangelsky, S. & Gamerro, J. C. (1966a). Estudio palinológico de la Formación Baqueró (Cretácico), provincia de Santa Cruz. II. Ameghiniana, 4(6), 201–209.
Archangelsky, S. & Gamerro, J. C. (1966b). Estudio palinológico de la Formación Baqueró (Cretácico), provincia de Santa Cruz. III. Ameghiniana, 4(7), 229–236.
Archangelsky, S. & Gamerro, J. C. (1966c). Estudio palinológico de la Formación Baqueró (Cretácico), provincia de Santa Cruz. IV. Ameghiniana, 4(10), 363–372.
Archangelsky, S. & Villar de Seoane, L. (1992). Estudios palinológicos de la Formación Baqueró (Cretácico) provincia de Santa Cruz, Argentina. V. En Actas del VIII Simposio Argentino de Paleobotánica y Palinología (p. 23–27). Corrientes.
Archangelsky, S. & Villar de Seoane, L. (1994). Estudios palinológicos de la Formación Baqueró (Cretácico) provincia de Santa Cruz, Argentina. VI. Ameghiniana, 31, 41–53.
Archangelsky, S. & Villar de Seoane, L. (1996). Estudios palinológicos de la Formación Baqueró (Cretácico) provincia de Santa Cruz, Argentina. VII. Ameghiniana, 33, 307–313.
Archangelsky, S. & Villar de Seoane, L. (1998). Estudios palinológicos de la Formación Baqueró (Cretácico) provincia de Santa Cruz, Argentina. VIII. Ameghiniana, 35, 7–18.
Archangelsky, S. & Villar de Seoane, L. (2005). Estudios palinológicos del Grupo Baqueró (Cretácico Inferior), Provincia de Santa Cruz, Argentina. IX. Polen bisacado de Podocarpaceae. Revista Española de Paleontología, 20, 37–56.
Balme, B. E. (1995). Fossil in situ spores and pollen grains: An annotated catalogue. Review of Palaeobotany and Palynology, 87, 81–323. https://doi.org/10.1016/0034-6667(94)00111-5
Barreda, V. D., Cúneo, N. R., Wilf, P., Currano, E. D., Scasso, R. A. & Brinkhuis, H. (2012). Cretaceous/Paleogene Floral Turnover in Patagonia: Drop in Diversity, Low Extinction, and a Classopollis Spike. PLoS ONE, 7(12), e52455. https://doi.org/10.1371/journal.pone.0052455
Carrizo, H., Fernández, E., Rosa, A. & Silva, S. (2018). Opera Lilloana 51 (1) (2018): Atlas de plantas fósiles. Colección Paleobotánica de la Fundación Miguel Lillo. 10.30550/b.opl/2018.v51n1.
Carrizo, M. A., Del Fueyo, G. M. & Archangelsky, S. (2012). Morfología y anatomía de un helecho creciendo bajo condiciones de estrés en el Aptiano de Santa Cruz, Argentina. Ameghiniana, 48(4), 605–617.
Césari, S. N., Limarino, C. O., Llorens, M., Passalia, M. G., Loinaze, V. P. & Vera, E.I. (2011). High-precision late Aptian Pb/U age for the Punta del Barco Formation (Baqueró Group), Santa Cruz Province, Argentina. Journal of South American Earth Sciences, 31, 426–431.
Cladera, G., Andreis, R., Archangelsky, S. & Cúneo, R. (2002). Estratigrafía del Grupo Baqueró, Patagonia (Provincia de Santa Cruz, Argentina). Ameghiniana, 39, 3–20.
Del Fueyo, G. M. & Archangelsky, A. (2002). Araucaria grandifolia Feruglio from the Lower Cretaceous of Patagonia, Argentina. Cretaceous Research, 23, 265-277.
Del Fueyo, G. M. & Archangelsky, S. (2005). A new araucarian pollen cone with in situ Cyclusphaera Elsik from the Aptian of Patagonia, Argentina. Cretaceous Research, 26, 757-768.
Del Fueyo, G. M., Villar de Seoane, L., Archangelsky, A., Guler, V., Llorens, M., Archangelsky, S., Gamerro, J. C., Musacchio, E. A., Passalia, M. G. & Barreda, V. D. (2007). Biodiversidad de las paleofloras de Patagonia Austral durante el Cretácico Inferior. Ameghiniana, Publicación Especial, 11, 101–122.
Herbst, R. (1960). Descripción de algunos ejemplares fértiles de Hausmannia (Protorhipis) papilio Fer., procedentes de los estratos de Baqueró, Santa Cruz. Acta Geologica Lilloana, 3, 227–232.
Herbst, R. (1962). Sobre las especies de Gleichenites de los sedimentos baqueroenses de Santa Cruz, Patagonia. Ameghiniana, 2, 141–151.
Herbst, R. (1966). Nuevos elementos de la flora fósil de la Formación Baqueró, Santa Cruz, Patagonia. Acta Geologica Lilloana, VIII, 75–89.
Herbst, R. (1971). Palaeophytologia Kurtziana. III. 7. Revisión de las especies argentinas del género Cladophlebis. Ameghiniana, 8(3–4), 265–281.
Kramer, K. U. & Green, P. S. (Eds.). (1990). Pteridophytes and Gymnosperms. Springer.
Korall, P., Conant, D. S., Metzgar, J. S., Schneider, H., & Pryer, K. M. (2007). A molecular phylogeny of scaly tree ferns (Cyatheaceae). American Journal of Botany, 94, 873–886.
Lafuente Díaz, M. A., Del Fueyo, G. M., D’Angelo, J. A. & Carrizo, M. A. (2021). Preserved chemistry of Cretaceous gymnosperm leaves in volcanic-ash deposits. Baqueró Group, Patagonia, Argentina. Cretaceous Research, 118.
Limarino, C. O., Passalia, M. G., Llorens, M., Vera, E. I., Perez Loinaze, V. S. & Césari, S. N. (2012). Depositional environments and vegetation of Aptian sequences affected by volcanism in Patagonia. Palaeogeography, Palaeoclimatology, Palaeoecology, 323–325, 22–41.
Llorens, M. (2003). Granos de polen de angiospermas de la Formación Punta del Barco (Aptiano), provincia de Santa Cruz, Argentina. Revista del Museo Argentino de Ciencias Naturales nueva serie, 5, 235–240.
Llorens, M. (2008a). Estudio palinológico de la Formación Punta del Barco, Cretácico Inferior (Aptiano) de la provincia de Santa Cruz, Argentina: Descripciones sistemáticas I: Esporas monoletes y triletes lisas y cinguladas. Ameghiniana, 45, 153–166.
Llorens, M. (2008b). Estudio palinológico de la Formación Punta del Barco, Cretácico Inferior (Aptiano) de la provincia de Santa Cruz, Argentina: Descripciones sistemáticas II: Esporas triletes apiculadas, murornadas, tricrasadas y perinadas. Implicancias paleoecológicas. Ameghiniana, 45, 273–288.
Llorens, M. (2012). Estudio palinológico de la Formación Punta del Barco (Aptiano) de la provincia de Santa Cruz, Argentina: Granos de polen de gimnospermas y caracterización de la flora. Ameghiniana, 49, 409–426. https://doi.org/10.5710/AMGH.29.06.2012.573
Llorens, M. & Perez Loinaze, V. S. (2016). Late Aptian angiosperm pollen grains from Patagonia: Earliest steps in flowering plant evolution at middle latitudes in southern South America. Cretaceous Research, 57, 66–78. https://doi.org/10.1016/j.cretres.2015.06.004
Llorens, M., Perez Loinaze, V. S., Passalia, M. G. & Vera, E. I. (2020). Palynological, megafloral and mesofossil record from the Bajo Grande area (Anfiteatro de Ticó Formation, Baqueró Group, Upper Aptian), Patagonia, Argentina. Review of Palaeobotany and Palynology, 273, 104137. https://doi.org/10.1016/j.revpalbo.2019.104137
Machado, M. A., Passalia, M. G., Vera, E. I. & Yañez, A. (2023). Ferns from the Arroyo Chacay flora (Huitrera Formation, Eocene), Río Negro Province, Argentina. Review of Palaeobotany and Palynology, 313, 104892.
Machado, M. A., Vera, E. I., Passalia, M. G. & Ponce, M. M. (2021). Eupolypod ferns with dryopteroid/thelypteroid traits from Arroyo Chacay (Huitrera Formation, Eocene), Río Negro Province, Argentina. Review of Palaeobotany and Palynology, 287, 104381.
Miller, I. M. & Hickey, L. J. (2008). The fossil flora of the Winthrop Formation (Albian–Early Cretaceous) of Washington State, USA. Part I: Bryophyta and Pteridophytina. Bulletin of the Peabody Museum of Natural History, 49, 135–180.
Nagalingum, N. S. & Cantrill, D.J. (2006). Early Cretaceous Gleicheniaceae and Matoniaceae (Gleicheniales) from Alexander Island, Antarctica. Review of Palaeobotany and Palynology, 138, 73–93. https://doi.org/10.1016/j.revpalbo.2005.11.001
Nishida, H., Uemura, K., Terada, K., Asakawa, T., Yabe, A., Yamada, T., Kurita, Y., Hinojosa, L.F. & Rancusi, M. (2014). Permineralized Matoniaceae fern from the Eocene Cranilahue Formation of Cocholgue, Chile, 4 International Palaeontological Congress, Abstract 580.
Panza, J. L., Marín, G., & Zubia, M. (1998). Hoja Geológica 4969-I, Gobernador Gregores, escala 1:250:000, provincia de Santa Cruz. Servicio Geológico Nacional, Boletín 239.
Passalia, M. G., Llorens, M., Césari, S. N., Limarino, C. O., Perez Loinaze, V. S., & Vera, E. I. (2016). Revised stratigraphic framework of the Cretaceous in the Bajo Grande area (Argentinean Patagonia) inferred from new U–Pb ages and palynology. Cretaceous Research, 60, 152–166.
Passalía, M. G., Llorens, M., Perez Loinaze, V., & Vera, E. I. (2017). Basal eudicot leaves from the latest Aptian–earliest Albian? of Patagonia. In Reunión de Comunicaciones de la Asociación Paleontológica Argentina, Libro de resúmenes (p. 60–61).
Passalia, M.G., Iglesias, A., Varela, A.N., Santamarina, P., Poiré, D.G. & Richiano, S.M. (2018). The fern Konijnenburgia alata in the mid-Cretaceous of Patagonia, and the Matoniaceae fossil record. Cretaceous Research, 89, 264–278. https://doi.org/10.1016/j.cretres.2018.03.026
Perez Loinaze, V. S., & Llorens, M. (2018). Palynology of the Baqueró Group (upper Aptian), Patagonia Argentina: An integrated study. Cretaceous Research, 86, 219–237.
Perez Loinaze, V. S., Vera, E. I., Passalia, M. G., Llorens, M., Friedman, R., Limarino, C. O., & Césari, S. N. (2013). High-precision U–Pb zircon age from the Anfiteatro de Ticó Formation: Implications for the timing of the early angiosperm diversification in Patagonia. Journal of South American Earth Sciences, 48, 97–105.
Pons, D. (1988). Le Mésozoïque de Colombie: macroflores et microflores. CNRS, Paris.
Sajjadi, F., & Playford, G. (2002). Systematic and stratigraphic palynology of Late Jurassic-earliest Cretaceous strata of the Eromanga Basin, Queensland, Australia: Part two. Palaeontographica Abteilung B, 261, 99–165.
Schrank, E. (2010). Pollen and spores from the Tendaguru Beds, Upper Jurassic and Lower Cretaceous of southeast Tanzania: Palynostratigraphical and paleoecological implications. Palynology, 34, 3–42. https://doi.org/10.1080/01916121003675710
van Konijnenburg-van Cittert, J. H. A. (1993). A review of the Matoniaceae based on in situ spores. Review of Palaeobotany and Palynology, 78, 235–267.
Vera, E. I., & Césari, S. N. (2012). Fossil woods (Coniferales) from the Baqueró Group (Aptian), Santa Cruz Province, Argentina. Anais da Academia Brasileira de Ciencias, 84, 617–625. https://doi.org/10.1590/S0001-37652012005000021
Vera, E. I., & Césari, S. N. (2015). New species of conifer wood from the Baqueró Group (Early Cretaceous) of Patagonia. Ameghiniana, 52, 468–471.
Vera, E. I., & Perez Loinaze, V. S. (2022). Ecological interactions in conifers (Agathoxylon and Protocupressinoxylon) from the Punta del Barco Formation (Baqueró Group, upper Aptian), Patagonia, Argentina. Cretaceous Research, 129, 105035. https://doi.org/10.1016/j.cretres.2021.105035
Vera, E., & Passalia, M. G. (2013). Gleicheniaceaephyllum san-martini, a new name for Gleichenites san-martini Halle emend. Herbst 1962. Ameghiniana
Vera, E. I., & Passalia, M. G. (2012). Korallipteris, a new genus for Mesozoic Gleichenia-like fern fronds. Geobios, 45, 421–428.
Villar De Seoane, L. M. (1999). Estudio comparado de Cyathea cyathifolia comb. nov. del Cretácico Inferior de Patagonia, Argentina. Spanish Journal of Palaeontology, 14, 157.
Villar De Seoane, L. M. (1996). Estudio morfológico de helechos fértiles hallados en la Formación Baqueró (Cretácico Inferior), Provincia de Santa Cruz, Argentina. Spanish Journal of Palaeontology, 11, 83–90.
Villar de Seoane, L. (2014). Estudio morfológico del género Classopollis (Pflug) Pocock & Jansonius (Cretácico, Argentina). Revista Brasileira de Paleontologia, 17(1), 91–104.
Villar de Seoane, L., & Archangelsky, S. (2014). Estudios palinológicos del Grupo Baqueró (Cretácico Inferior), provincia de Santa Cruz, Argentina. X. Polen de Gymnospermae y Apéndice Final. Revista del Museo Argentino de Ciencias Naturales nueva serie, 16(1), 33–44.