Articulo original
Original article
35 (2) | Jul - Dic, 2025
Facultad de Ciencias Exactas y Naturales y Agrimensura (UNNE)
Open Access: https://revistas.unne.edu.ar/index.php/fce
E-mail: revistafacena@exa.unne.edu.ar
Tres taxones nuevos para la Xiloflora del Mioceno de la Formación San José, Tucumán, Argentina
Three new taxa for the Miocene Xyloflora of the San José Formation, Tucumán, Argentina
Baez, Johanna * 1 , Crisafulli, Alexandra 2 & Martinez, Ramona Mercedes 2
1. Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria Paraná, Oro Verde, Entre Ríos, Argentina.
2. Facultad de Ciencias Exactas y Naturales y Agrimensura (FaCENA-UNNE). Centro de Ecología Aplicada del Litoral (CONICET-UNNE). Corrientes, Argentina.
* Autor de correspodencia: johannasbaez@gmail.com
Recibido/Received: 29 de Abril, 2025 | Aceptado/Acepted: 24 de Agosto, 2025 | Publicado/Published: 6 de Febrero, 2026.
Como citar este artículo: Baez, J., Crisafulli, A. & Martinez, R. M. (2025). Tres taxones nuevos para la Xiloflora del Mioceno de la Formación San José, Tucumán, Argentina. Revista FACENA 35(2), 223-253. Doi: https://doi.org/10.30972/fac.3528844
Resumen: El presente trabajo da a conocer dos nuevos géneros y tres especies nuevas de maderas fósiles asignadas a angiospermas dicotiledóneas, procedentes de la localidad de Tiu Punco donde aflora la Formación San José (Mioceno medio), en el valle de Santa María, provincia de Tucumán, Argentina. Las muestras, recolectadas en niveles fluvio-lacustres, fueron analizadas anatómicamente y son afines a las Fabaceae, Bignoniaceae y Combretaceae. Estos hallazgos aportan evidencia clave sobre la composición florística y las condiciones ecológicas de los ecosistemas miocénicos del noroeste argentino. A su vez, se discuten sus implicancias paleoambientales y su relevancia en el Grupo Santa María, destacando la continuidad y diversidad de la cubierta vegetal en esta secuencia geológica. El estudio contribuye a la comprensión de la evolución ecoanatómica de la vegetación neógena en Sudamérica y fortalece el registro paleobotánico regional.
Palabras claves: Formación San José; Mioceno medio; Leños Fósiles; Fabaceae; Bignoniaceae, Combretaceae.
Abstract: This paper describes two new genera and three new species of fossil wood assigned to dicotyledonous angiosperms, from the locality of Tiu Punco, where the San José Formation (middle Miocene) outcrops in the Santa María Valley, Tucumán Province, Argentina. The samples, collected from fluvial-lacustrine levels, were anatomically analysed and are related to the Fabaceae, Bignoniaceae and Combretaceae. These findings provide key evidence regarding the floristic composition and ecological conditions of Miocene ecosystems in north-western Argentina. Their palaeoenvironmental implications and relevance to the Santa María Group are discussed, highlighting the continuity and diversity of the vegetation cover in this geological sequence. This study contributes to our understanding of the eco-anatomical evolution of Neogene vegetation in South America and strengthens the regional palaeobotanical record.
Keywords: San José Formation; middle Miocene; Fossil Woods; Fabaceae; Bignoniaceae; Combretaceae.
Introducción
En esta contribución se describen dos nuevos géneros y tres especies de maderas fósiles afines a Fabaceae, Bignoniaceae y Combretaceae, colectadas por Herbst en la década de 1990 durante una de sus numerosas campañas en la localidad de Tiu Punco, donde aflora la Formación San José, unidad que forma parte del Grupo Santa María, provincia de Tucumán, Argentina (Figura 1), caracterizada por niveles bioclásticos que han sido objeto de estudios tafonómicos enfocados en parámetros bioestratinómicos como orientación, desarticulación y fragmentación (Benítez, 2022).
La edad de esta Formación es Mioceno medio, ~ 12 Ma, (Gavriloff y Bossi, 1992; Bossi et al. 1997; Gavriloff, 1999; Kleinert y Strecker, 2001; Morton y Herbst, 2003) de acuerdo con estudios radiométricos y paleontológicos. Los aportes de Anzótegui (1997, 2001, 2002), García y Herbst (1997), Anzótegui y Cristalli (2000), Anzótegui y Morton (2000) Anzótegui y Ferraro (2000), Herbst et al. (2000), Mautino y Anzótegui (2000) Mautino et al. (2004), Morton y Herbst (2003), Anzótegui y Herbst (2004), Mautino (2007, 2010, 2011); Mautino y Anzótegui (2014), Robledo (2017), Robledo et al. (2018), Baez et al. (2019b), Anzótegui et al. (2019), Baez et al. (2021) brindaron información referente a los fósiles presentes en la secuencia de la porción inferior del Grupo Santa María, lo que permitió observar variedad de estructuras y proveyó herramientas de comparación con otras unidades de la misma edad.
La tafoflora está compuesta por Malvaciphyllum quenquiadensis Anzótegui y Cristalli 2000 (Malvaceae) y Myrciophyllum sp. (Myrtaceae) (Anzótegui, 2002), Schinopsis aff. lorentzii (Griseb) Eng. (Anacardiaceae), Ziziphus sp. (Rhamnaceae), Prosopis sp., Erythrina dominguezii, Erythrina aff. dominguezii Hassl (Fabaceae). Dentro de los frutos encontrados, pueden mencionarse Cyperocarpus sp.1 y C. sp.2 (Cyperaceae), Tibouchina sp. (Melastomataceae), Baccharis sp. y Asteracarpus gomez-belloi (Asteraceae) (Anzótegui y Herbst, 2004). Los carófitos Chara strobilocarpa Reid y Groves y Chara sp. provienen de los niveles fangolíticos, a veces calcáreos, que representan condiciones lacustres (García y Herbst, 1997).
Además, se ha identificado en esta secuencia a Mimosoxylon santamariensis (Fabaceae) (Lutz, 1987) y Baez et al. (2019b) describieron un leño de Combretaceae.
Baez (2022) señaló la ocurrencia de numerosas especies de maderas de angiospermas y otras novedades para la xilotafología de las formaciones que integran este Grupo.
La paleofauna de invertebrados hallada está constituida por moluscos y ostrácodos, es conocida a través de los numerosos trabajos sintetizados en Herbst et al. (2000) y Morton y Herbst (2003, 2012), cuya preservación y disposición han sido analizadas desde una perspectiva bioestratinómica, revelando distintos grados de retrabajo y acción hidrodinámica (Benítez, 2022).
La gran mayoría de vertebrados están representados por mamíferos incluyendo notongulados, xenartros y rodéntidos (Herbst et al. 2000). Hay escasos restos de aves y placas aisladas de tortugas (Herbst et al. 2000) y pequeños peces de la familia Anablepidae (Anzótegui y Herbst, 2004).
El interés que despiertan los nuevos registros del presente trabajo motiva a continuar con los estudios de determinación sistemática de los leños y de la paleoflora en general que tantas riquezas aún siguen revelando a la vez que se enmarca como un homenaje a los investigadores que contribuyeron significativamente al estudio de esta región.
marco geológico
Formación San José
Constituye la unidad basal del Grupo Santa María que fue definida por Galván y Ruíz Huidobro (1965). Se compone de areniscas, limos y arcillitas, amarillo verdoso predominantemente, poco cementados y erosionados. Muestran un típico paisaje de badlands. Hay otros sedimentos constituidos por pelitas margosas, calizas y arcillitas foliosas y en menor proporción algunos conglomerados (Figura 2).
Bossi y Palma (1982) señalaron que en la localidad tipo (San José, valle de Santa María) tiene una potencia de 250 m, pero aumenta sostenidamente hacia al norte hasta alcanzar cerca de los 1000 m en la quebrada Agua Negra, Salta.
La Formación San José es interpretada como una unidad fluvio-lacustre, con cuerpos de agua (lagos), aunque parte de la secuencia indica condiciones palustres. Estas inferencias están sustentadas por las características sedimentológicas y el registro paleontológico (Herbst, 2016).
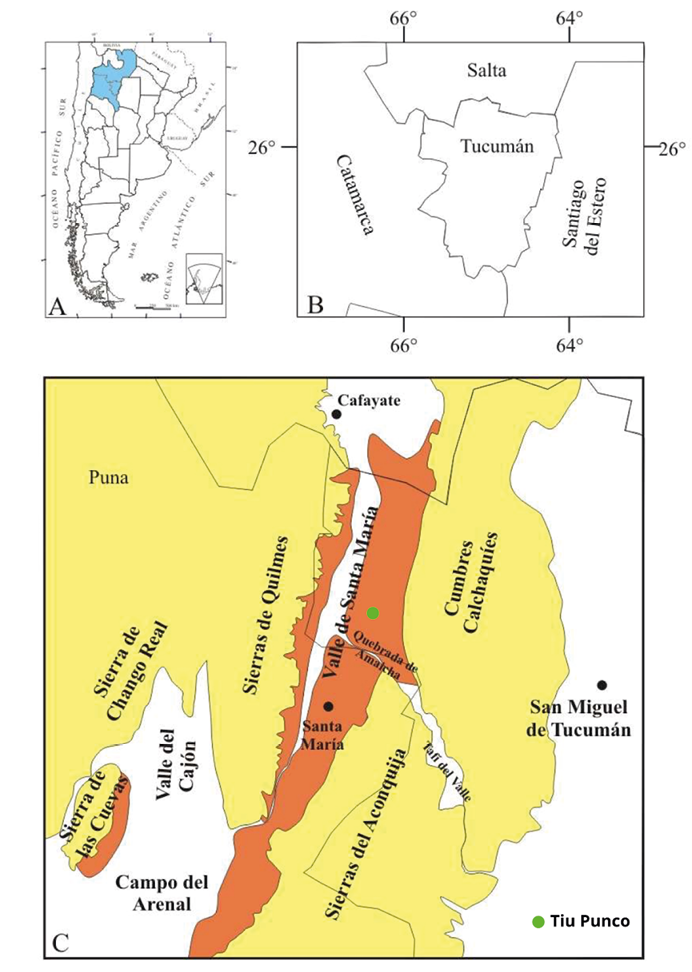
Figura 1. Ubicación del área donde aflora el Grupo Santa María (representada en color marrón) y localización de Tiu Punco en proximidad a la Quebrada de Amaicha del Valle. Tomado y modificado de Moyano (2009).
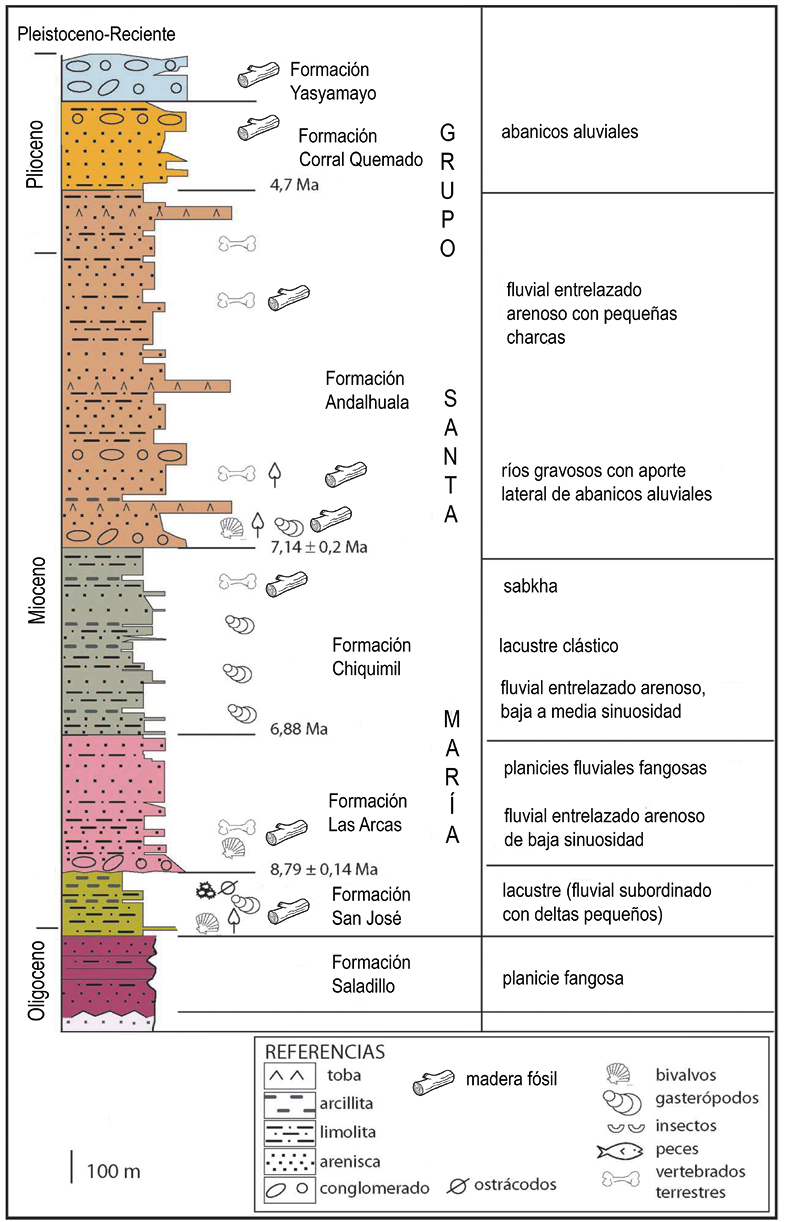
Figura 2. Columna estratigráfica de las formaciones que integran el Grupo Santa María. Tomado y modificado de Georgieff (2014).
MATERIALES Y MÉTODOS
Los leños descriptos en este aporte son ejemplares permineralizados por silicificación, que han preservado sólo la anatomía del xilema secundario. Para su análisis se realizaron cortes transversales, longitudinales radial y tangencial, respectivamente y pulidos al punto óptimo de delgadez (40 μm) de acuerdo con la metodología propuesta por Hass y Rowe (1999). Las observaciones se realizaron con microscopio óptico Leica IC C 50 y lupa estereoscópica Leica M50, cámara digital Leica EC3. Para las medidas de los distintos elementos anatómicos se registraron promedios de 25 mediciones, en todos los casos se cita primero la media y entre paréntesis las medidas mínimas y máximas.
La terminología empleada se basa en el IAWA List of Microscopic Features for Hardwood Identification (1989), Carlquist (2001) y las bases taxonómicas de Zuloaga y Morrone (1999), The World Flora Online (2020) y POWO (2023). Se siguió las clasificaciones sistemáticas de Chase et al. (2016) y Nassim et al. (2017). Asimismo, se consultaron bases de datos en línea como Inside Wood (2004–en adelante); glosarios especializados (Cozzo, 1964), clasificaciones estándar de Chattaway (1932), recopilaciones bibliográficas de Gregory (1994) y el catálogo de Gregory et al. (2009) sobre los géneros y especies fósiles de Magnoliopsidas.
De acuerdo con Brea et al. (2010) y Avise y Johns (1999), se sigue con la propuesta de determinar con nombres genéricos fósiles a los ejemplares que sólo preservan el xilema secundario, a pesar de la sugerencia de Collinson (1986) de que se use géneros actuales para taxones a partir del Plioceno.
Los anillos de crecimiento, porosidad, números de vasos por mm², diámetro, longitud y tipo de los vasos, tipo de placa de perforación, tipo de parénquima axial, tipo de punteaduras, cristales y tilosis son caracteres anatómicos que permiten ser relacionados con factores climáticos y condiciones ecológicas, principalmente los que participan en la conducción y seguridad en el transporte de agua (Wheeler y Baas, 1991; Lindorf, 1994; Moglia y Giménez, 1998; Ewers et al. 2023); es por ello que se los analizó para completar el estudio paleoecológico de esta unidad estratigráfica.
Carlquist (1977) estableció dos índices de los vasos del xilema secundario para cuantificar el grado de mesomorfía o xeromorfía exhibida por las plantas que relacionan la anatomía de la madera con la humedad ambiental donde crecieron las mismas. El uso de estos índices ayuda a comprender las estrategias hidráulicas de las plantas (Ewers et al. 2023). La densidad de los vasos está presumiblemente relacionada con la seguridad hidráulica a través de la redundancia y la resistencia a la embolia y es una característica esencial de las maderas xeromorfas que tienden a tener muchos vasos estrechos. El índice de Vulnerabilidad (IV) es igual al cociente entre el diámetro promedio de los elementos de los vasos y la densidad de vasos por mm². Para un valor mayor a 1 son vulnerables a la cavitación y menor a 1 son seguros en la conducción del agua. Generalmente, cuanto más alto sea el valor del diámetro de vasos menor será su densidad por mm² y más grande será el valor de IV (Moglia y Giménez, 1998).
El Índice de Mesomorfía (IM) se obtiene multiplicando el Índice de Vulnerabilidad por la longitud promedio de los elementos de los vasos. Un valor mayor a 200 significa que las maderas son mesomórficas (IV > 1; IM> 200) y valores por debajo de 200 corresponden a maderas xeromórficas (IV < 1; IM < 200) (Carlquist 1977, 2001).
Repositorio
Los materiales analizados son fragmentos de maderas pertenecientes a la Colección Paleontológica de la UNNE, Dr. Rafael Herbst, Sección Paleobotánica (bajo el acrónimo CTES- PB) y a la sección Preparados Micropaleontológicos (CTES-PMP), de Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste, ubicada en el Centro de Ecología Aplicada del Litoral, (CECOAL-CONICET/UNNE), Corrientes, Argentina. El material restante pertenece a la Colección Paleobotánica (bajo el acrónimo: LIL-PB) de la Fundación Miguel Lillo, Tucumán, Argentina.
resultados
Sistemática Paleobotánica
Eudicotyledoneae sensu Chase et al. 2016
Orden Fabales Chase et al. 2016
Familia Fabaceae Juss 1789 ̶ Leguminosae Nassim et al. 2017
Tiopuncoxylon gen. nov.
Especie tipo. Tiopuncoxylon angiona sp. nov.
Derivatio nominis. El género alude a la localidad de origen del material.
Diagnosis. Diffuse porosity. Solitary and multiple radial vessels of two or more elements with sui generis ovoid shape. Perforation plates exclusively simple. Intervessel pits opposite, alternate, bordered and vestured. Apotracheal axial parenchyma diffuse and paratracheal parenchyma vasicentric, confluent in narrow bands. Homocellular rays composed of procumbent cells. Rays multiseriate and triseriate; fibres nonseptate.
Diagnosis. Porosidad difusa. Vasos solitarios y múltiples radiales de dos a más elementos con forma ovoidal sui generis. Placas de perforación exclusivamente simples. Punteaduras intervasculares opuestas, alternas, areoladas y ornadas. Parénquima axial apotraqueal difuso y paratraqueal vasicéntrico, confluente en bandas angostas. Cristales prismáticos en células del parénquima axial con cámaras. Radios homocelulares compuestos por células procumbentes. Radios multiseriados y triseriados; fibras no septadas.
Tiopuncoxylon angiona sp. nov.
Figura 3
Diagnosis. Como la del género por monotipia.
Derivatio nominis. El epíteto específico hace referencia al vaso (angion). Los mismos tienen una fisonomía peculiar por la diferencia entre el diámetro radial con respecto al tangencial.
Holotipo. CTES PB 14710,CTES-PMP: 3786 a, b, c
Localidad. Tiu Punco.
Procedencia geográfica. Tucumán, Argentina.
Procedencia estratigráfica. Formación San José.
Edad. Mioceno medio.
DESCRIPCIÓN: los anillos de crecimiento son distinguibles. La porosidad es difusa. Los vasos son solitarios (40%), múltiples radiales y tangenciales de 2 (30%), 3 (10%) y más elementos (20%) (Figura 3. 1–4). Tienen un contorno ovoidal y disposiciones peculiares no vistos frecuentemente. Los vasos tienen un diámetro tangencial de 86 (31–158) μm y un diámetro radial de 329 (158-573) μm. Las paredes con un espesor de 19 (6,3 – 31) μm; la densidad de los vasos es de 9 (6–1) por mm². Vasos con contenido y aparente presencia de sustancias tóxicas fenólicas (sensu Schweingruber et al. 2007) (Figura 3. 5-9). El parénquima axial es apotraqueal difuso (Figura 3. 3) y paratraqueal vasicéntrico (Figura 3. 9), confluente (Figura 3.7). Las fibras son abundantes, poseen contorno poligonal, con un diámetro promedio de 25 (19–31) μm y espesor de sus paredes de 8 μm (Figura 3. 10). Los radios presentan trayecto recto. Los radios son homocelulares compuestos por células procumbentes (Figura 3. 11–13), se observa el parénquima septado de 13 células de alto (Figura 3. 14). Las punteaduras radiovasculares son alternas y unas pocas opuestas y ornadas (Figura 3.15). El diámetro es de 17 (13–20) μm, las placas de perforación son simples. En las fibras se observan punteaduras simples. Los elementos de vasos tienen una longitud de 129 (87-200) μm (Figura 3. 16). Los radios son multiseriados (70%) y triseriados (30%) (Figura 3. 16, 17). Presentan una altura de 27 (21–33) células. La densidad es de 23 radios por mm². Las punteaduras intervasculares son alternas y unas pocas opuestas y ornadas (Figura 3. 18). Las fibras no son septadas. Hay cristales prismáticos en cámaras de las células del parénquima axial (Figura 3. 19, 20). El índice de vulnerabilidad fue de 9.56; y el IM dio un valor de 1233,24.
DISCUSIÓN: El análisis de las características anatómicas de este fosilitaxón presenta la siguiente combinación: porosidad difusa, vasos de contorno ovoidal con marcada diferencia en el diámetro radial con respecto al tangencial; parénquima axial apotraqueal difuso y paratraqueal vasicéntrico a confluente; cristales en cámaras de las células del parénquima axial, placas de perforación simples, punteaduras intervasculares y radiovasculares alternas, algunas opuestas y ornadas, fibras no septadas y radios homocelulares compuestos por células procumbentes, multiseriados y triseriados. Esta combinación de caracteres se corresponde con los rasgos anatómicos típicos de las fabáceas, tal como ha sido documentado en estudios previos sobre maderas fósiles y actuales (Pujana et al., 2011; Moya, 2018; Gregory et al., 2009).
Los caracteres como la porosidad difusa, tipo de vasos, el parénquima apotraqueal difuso, radios multiseriados son compartidos con géneros de la familia Malvaceae (Ramos et al. 2017; Pujana, 2022). Sin embargo, la presencia de placas de perforación escalariformes y punteaduras escalariformes en algunos taxones de esta familia las diferencias del ejemplar analizado.
Con ejemplares de Moraceae, comparte los tipos de vasos, las placas de perforación simples, fibras abundantes, parénquima paratraqueal vasicéntrico, bandeado o confluente, parénquima apotraqueal difuso y los radios multiseriados. Pero, difiere en presentar radios uniseriados y cristales prismáticos en las células de los radios, punteaduras alternas y no ornadas.
Otra familia de angiospermas que comparte la mayoría de los rasgos anatómicos es Combretaceae (Metcalfe y Chalk, 1950), pero se aparta por tener radios uni y biseriados en la totalidad de sus especies.
De esta comparación se desprende que este ejemplar muestra mayor afinidad con Fabaceae. Se establecieron comparaciones con las siguientes maderas fósiles de esta familia, por ejemplo, Peltophoroxylon (Ramanujan) Müller-Stoll y Mädel, Paracacioxylon Müller-Stoll y Mädel, Paraalbizioxylon (Gros) emend. Martínez, Menendoxylon Lutz, Dichrostachyoxylon Müller Stoll y Mädel (ver tablas en Brea et al. 2010; Martínez, 2010; Franco y Brea, 2013) y Gleditsioxylon Müller-Stoll y Mädel.
Peltophoroxylon variegatum (Ramanujan) Müller-Stoll y Mädel del Mio-Plioceno de la India se distingue por tener punteaduras alternas, fibras septadas, parénquima vasicéntrico, aliforme, a veces confluente y con radios heterocelulares en algunos sectores.
Paracacioxylon se aparta por presentar porosidad semi-circular a difusa y agrupados “clusters”, parénquima vasicéntrico escaso a raro y unilateral y radios uniseriados. Paraalbizioxylon se distingue por poseer porosidad semi-circular a difusa, vasos solitarios en un 70% y agrupados, radios uniseriados y algunos parcialmente biseriados, ausencia de parénquima apotraqueal difuso. Dichrostachyoxylon Müller-Stoll y Mädel del Oligoceno-Mioceno de Rumania se diferencia por presentar un parénquima axial predominantemente vasicéntrico estrecho, ocasionalmente aliforme y raramente confluente, además de vasos agrupados en pequeños grupos radiales. En cuanto a los radios, el género exhibe estructuras homogéneas o ligeramente heterogéneas, con un ancho de 1 a 10 células, mientras que la especie aquí descripta presenta radios multiseriados y triseriados.
Gleditsioxylon Müller-Stoll y Mädel se diferencia por el tipo de porosidad, la presencia de vasos agrupados, el parénquima vasicéntrico con cristales, radios uniseriados, parcialmente biseriados y engrosamientos helicoidales (Cuadro 1 en Martínez y Rodriguez-Brizuela 2011). Amerita asignar este leño a la familia Fabaceae por la variedad de los rasgos que presenta, pero se propone la creación de un nuevo género y especie por la combinación de los elementos anatómicos distintivos que presenta.
Distribución estratigráfica del taxón en el Neógeno de América del Sur. Tiu Punco (Tucumán-Argentina).
Afinidad botánica. Fabaceae.
Distribución actual de la Familia. Cosmopolita. Regiones tropicales.

Figura 3. Tiopuncoxylon angiona. CTES-PMP: 3786 a, b, c. a. Corte Transversal (1–10). 1. Aspecto general mostrando la porosidad difusa. 2–4. Tipos de vasos y anillos de crecimiento distinguible. 3–9 Detalle de los vasos con contenido y sustancias fenólicas. 3. Parénquima apotraqueal difuso. 7, 9. Parénquima paratraqueal vasicéntrico a confluente. 10. Detalle de las fibras de contorno poligonal. b. Corte longitudinal radial (11–15). 11–13. Radios homocelulares. 14. Detalle del parénquima septado. 15. Punteaduras radiovasculares alternas, algunas opuestas y ornadas. c. Corte longitudinal tangencial (16–20). 16–17. Distribución y frecuencia de los radios. 18. Punteaduras intervasculares alternas, unas pocas opuestas y ornadas. 19–20. Cristales prismáticos en cámaras de las células del parénquima axial. Escalas: 1, 11, 16 = 200 μm; 2–4= 50 μm; 5–8, 12, 13, 17, 18= 25 μm; 9, 10, 14, 15, 19, 20= 12 μm.
Orden Lamiales Bromhead 1840
Familia Bignoniaceae Juss.1789
Tabebuiaxylon gen. nov.
Especie tipo. Tabebuiaxylon eorigida sp. nov.
Derivatio nominis. El nombre genérico alude a su parecido con el género actual Tabebuia.
Diagnosis. Growth rings distinct. Porosity semicircular to diffuse. Vessels solitary, radial multiples of two or more elements and in clusters. Perforation plates exclusively single. Intervessel and opposite-ray vessel pits, small, less than 4 μm. Paratracheal axial parenchyma vasicentric, confluent and aliform. Tyloses and septate parenchyma present with or without crystals. Homocellular rays composed exclusively of square cells. Rays uniseriate, partially biseriate, some biseriate and triseriate; storied; with straight bands of parenchyma cells between them; septate and nonseptate fibres.
Diagnosis. Anillos de crecimiento distintos. Porosidad semicircular a difusa. Vasos solitarios, múltiples radiales de dos a más elementos y en clusters. Placas de perforación exclusivamente simples. Punteaduras intervasculares y radiovasculares opuestas, pequeñas, menores a 4 μm. Parénquima axial paratraqueal vasicéntrico, confluente y aliforme. Presencia de tilosis y parénquima septado con o sin cristales. Radios homocelulares compuestos exclusivamente por células cuadradas. Radios uniseriados, parcialmente biseriados, algunos biseriados y triseriados; estratificados, con bandas rectas de células parenquimáticas entre ellos; fibras septadas y no septadas.
Tabebuiaxylon eorigida sp. nov.
Figura 4
Diagnosis. Como la del género por monotipia.
Derivatio nominis. el epíteto específico alude a una especie ancestral a la actual Tabebuia rigida
Holotipo. CTES PB 14711, CTES-PMP: 3813 a, b, c.
Procedencia geográfica. Tucumán, Argentina
Localidad. Tiu Punco
Procedencia estratigráfica. Formación San José
Edad. Mioceno medio
DESCRIPCIÓN: Fragmento de leño silicificado que en corte transversal muestra los anillos de crecimiento ausentes. Presenta porosidad semi-circular a difusa. Los vasos son solitarios (10 %), múltiples radiales de 2 (30%), 3 (34%) y más elementos (11%) y agrupados (15%). Los vasos tienen un diámetro tangencial de 110 (60–180) μm y un diámetro radial de 165 (100–260) μm, las paredes con un espesor de 12 μm, la densidad de vasos es de 11 por mm². El parénquima axial es paratraqueal vasicéntrico, confluente y aliforme. Las fibras son abundantes, poligonales, con un diámetro de 12 μm y el espesor de sus paredes de 4 μm (Figura 4. 1–5). Los radios presentan un trayecto sinuoso a recto. Los radios son homocelulares, con células cuadradas. Los elementos de vasos tienen una longitud de 420 (100–630) μm, las placas de perforación son simples y las punteaduras radiovasculares son opuestas, pequeñas, menores a 4 μm (Figura 4. 6, 7). Los radios son uniseriados, uniseriados parcialmente biseriados, biseriados y triseriados. Las células que los componen son de contorno circular. Tienen una altura de 8 (4–10) células. La densidad es de 6 (4–10) radios por mm². Presentan estratificación. Fibras septadas escasas (Figura 4. 8, 9). El índice de vulnerabilidad registrado fue de 10, y el índice de mesomorfía alcanzó un valor de 4200.
DISCUSIÓN. Pace y Angyalossy (2013) agruparon a los integrantes de la familia Bignoniaceae por las siguientes características: porosidad circular, semi-circular a difusa; anillos de crecimiento distintos, generalmente marcados por parénquima marginal, parénquima paratraqueal escaso, vasicéntrico y aliforme; parénquima difuso presente o ausente. Los vasos son ovalados a circulares, solitarios, en múltiples de dos o tres, con placas de perforación simples; punteaduras intervasculares circulares y alternas, las punteaduras radiovasculares similares a las punteaduras intervasculares en tamaño y forma. La tilosis está presente, en forma escasa en las lianas. En las fibras libriformes, generalmente se ven punteaduras pequeñas y poco visibles. Los cristales son poco frecuentes y se distribuyen al azar en la familia; cuando están presentes suelen estar en las células de los radios y ocurren más raramente en el parénquima axial; pueden ser prismáticos, alargados o aciculares.
En las Bignoniaceae se pueden diferenciar tres grados de estructuras estratificadas: ausentes; presente para todas las células; y presente sólo para el parénquima axial y las fibras. La mayoría de los géneros de esta familia carecen completamente de estructura estratificada, considerado éste un carácter ancestral.
Por otra parte, hay cuatro tipos diferentes de radios: a) todos los radios son homocelulares, b) radios homocelulares y heterocelulares con células del cuerpo procumbentes y una fila de células marginales cuadradas, c) radios heterocelulares con células procumbentes y dos a cuatro células marginales erectas o cuadradas y d) radios heterocelulares con células procumbentes, cuadradas y erectas mezcladas.
Todos estos caracteres anatómicos tal vez, estén relacionados con la distribución tropical de la familia; las demás características de la madera son más variables. Consecuentemente con los rasgos descriptos más arriba, el leño analizado es una Bignoniaceae.
Dentro de las Bignoniaceae fósiles, se confrontó los caracteres del leño estudiado con Bignonioxylon americanum Moya y Brea 2018, de la Formación Arroyo Feliciano del Pleistoceno de Entre Ríos. Empero, éste se diferencia por presentar porosidad difusa, variedad en el parénquima aliforme (con alas cortas y unilateral), parénquima axial apotraqueal difuso, parénquima vasicéntrico septado, cristales, radios uniseriados y biseriados solamente y estratificación parcial e irregular de radios, fibras y/o parénquima. Asimismo, la muestra de leño de la Formación San José tiene una estrecha relación con el género actual Tabebuia. La más afín al material es: T. rigida Urban. La presencia de radios estratificados con dos bandas de células parenquimáticas entre ellos es la característica distintiva y comparte con esta madera fósil, por ello se propone la creación de un nuevo género.
También se comparó con Elizabethiaxylon patagonicum Ruíz, Pujana, Brea 2024 (Malvaceae); Tabebuiaxylon eorigida comparten ciertos caracteres, como la ausencia de anillos de crecimiento y la presencia de placas de perforación simples, presentan diferencias anatómicas sustanciales que impiden su inclusión dentro de un mismo taxon. Estas diferencias se evidencian en la disposición y densidad de los vasos, el tipo de parénquima axial y la estructura de los radios, lo que respalda su asignación a especies distintas.
A los fines comparativos, se cotejaron también los caracteres de Terminalioxylon lajaum Ramos, Brea y Kröhling 2012 (Combretaceae) y Piptadenioxylon chimeloi Suguio y Mussa 1978 (Fabaceae), con los que T. eorigida comparte varios rasgos anatómicos. Sin embargo, difiere en el ancho y la estratificación de los radios: T. lajaum presenta radios uniseriados, algunos parcialmente biseriados, homocelulares y constituidos por células procumbentes, mientras que P. chimeloi muestra radios mayormente triseriados y también homocelulares con células procumbentes. En contraste, el espécimen de Tiu Punco presenta radios uniseriados, parcialmente biseriados, biseriados y triseriados, con estructura estratificada y células cuadradas, lo que refuerza su distinción anatómica y taxonómica respecto a estas especies.
Distribución estratigráfica del taxón en el Neógeno de América del Sur. Formación San José (Tucumán), Argentina.
Afinidad botánica. Tabebuia rigida Urban.
Distribución actual. Zonas intertropicales de América extendidas desde México y el Caribe.
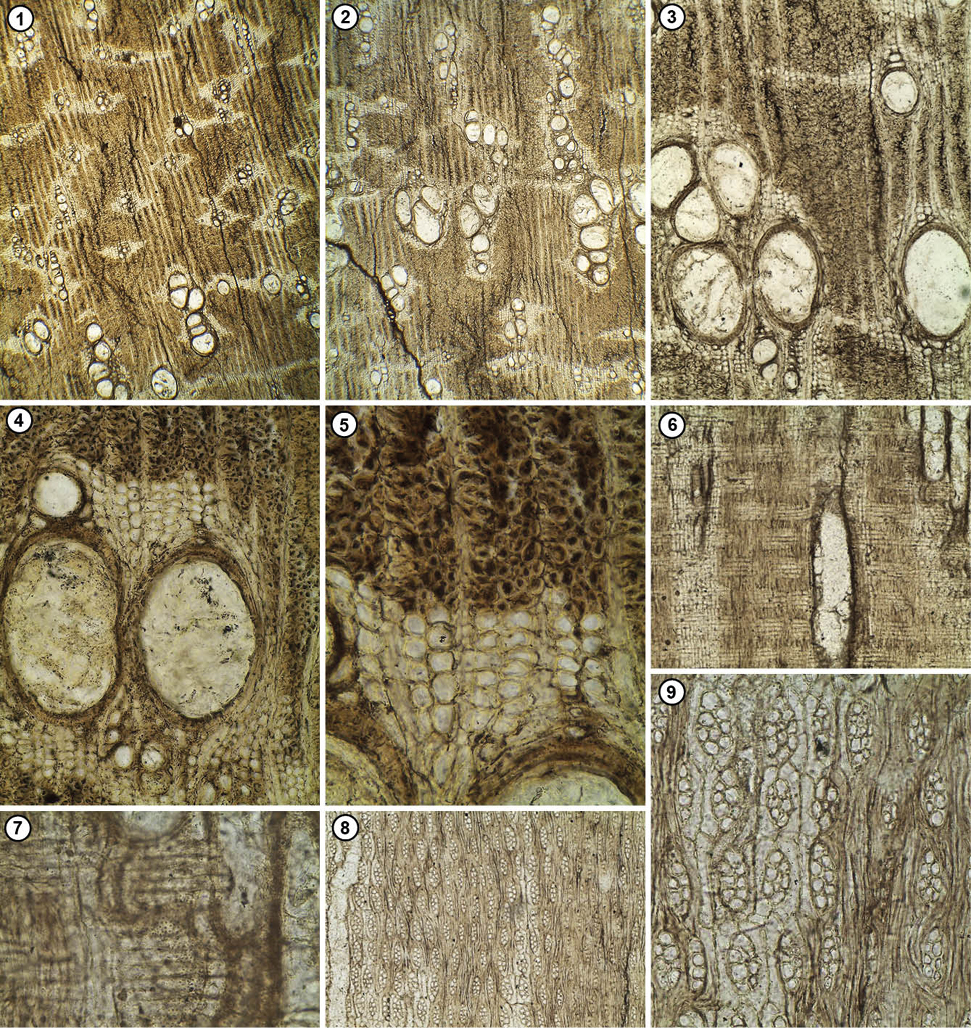
Figura 4. Tabebuiaxylon eorigida. CTES-PMP: 3813 a, b, c. a. Corte Transversal (1–5). Anillos de crecimiento marcados, porosidad semi-circular a difusa. Tipo de vasos. Parénquima paratraqueal vasicéntrico, confluente y aliforme. Fibras de contorno poligonal. b. Corte longitudinal radial 6, 7. Radios homocelulares con células cuadradas. Placas de perforación simples; en 7 punteaduras radiovasculares. c. Corte longitudinal tangencial 8, 9. Tipo y estratificación de los radios. Escalas: 1, 2, 6, 8= 200 μm; 3, 7, 9= 80 μm; 4, 5= 45 μm.
Orden Myrtales
Familia Combretaceae Brown 1810
Terminalioxylon (Schönfeld) Madel y Müller-Stoll 1973
Especie tipo. Terminalioxylon naranjo (Schönfeld) Madel y Müller-Stoll 1973
Terminalioxylon calchaquii sp. nov.
Figura 5
Derivatio nominis. La especie está dedicada a la memoria de Juan Calchaquí, valeroso Cacique que encabezó la resistencia nativa contra la dominación española siendo su centro de operaciones Tolombón.
Specific diagnosis. Growth rings indistinct. Diffuse porosity. Solitary, multiple short radial vessels, rarely 4. Perforation plates simple, intervessel pits alternate, spaced, ornate, oval. Paratracheal axial parenchyma vasicentric, confluent, tending to aliform. Polygonal fibres, nonseptate. Homocellular rays with uniseriate procumbent cells, with biseriate and biseriate portions, crystals in ray cells and axial parenchyma.
Diagnosis específica. Anillos de crecimiento indistintos. Porosidad difusa. Vasos solitarios, múltiples radiales cortos y raramente de 4. Placas de perforación simples, punteaduras intervasculares alternas, espaciadas, ornadas, ovales. Parénquima axial paratraqueal vasicéntrico, confluente con tendencia a aliforme. Fibras poligonales, no septadas. Radios homocelulares con células procumbentes uniseriados, con porciones biseriadas y biseriados. Cristales en las células de los radios y en el parénquima axial.
Holotipo. LIL-PB: 045-046-047
Localidad. Tiu Punco
Procedencia geográfica. Tucumán, Argentina.
Procedencia estratigráfica. Formación San José
Edad. Mioceno medio
DESCRIPCIÓN: Fragmento de leño que en corte transversal presenta anillos de crecimiento no marcados. La porosidad es difusa. Los vasos son solitarios (10%), múltiples radiales de 2 (65%), 3 (21%) y raramente de 4 (4%) de contorno ovoidal (Figura 5. 1). Los vasos tienen un diámetro tangencial de 80 (45–105) μm y un diámetro radial de 125 (80–176) μm, las paredes con un espesor de 5 μm, la densidad de vasos promedio es de 10 por mm². Vasos con contenido. El parénquima axial es paratraqueal vasicéntrico, confluente con tendencia a aliforme. Las fibras poseen contorno poligonal (Figura 5. 2–4). Los radios presentan trayecto sinuoso y son muy finos. Los radios son homocelulares compuestos por células procumbentes; tienen abundantes cristales distribuidos en las células de los radios y en las células del parénquima axial (Figura 5. 5–7). Los elementos de vasos tienen una longitud de 510 (500–700) μm y placas de perforación simples (Figura 5. 8). Las punteaduras intervasculares son alternas, espaciadas, ornadas y ovales. (Figura 5. 9). Los radios son uniseriados (50%), algunos de ellos presentan porciones biseriadas (30%) y unos pocos biseriados (20%). Presentan una altura promedio expresada en número de células de 16 (8–22). La densidad es de 9 radios por mm². Fibras no septadas. Presenta parénquima septado con cristales (Figura 5. 10). El índice de vulnerabilidad es de 8, y el índice de mesomorfia fue de 4080.
Discusión. El leño es asignado a las Combretaceae por presentar las características descriptas y principalmente por tener radios uniseriados y biseriados, distingue a las maderas de esta familia de las Fabaceae que poseen radios uniseriados a multiseriados.
El ejemplar comparte con el género Terminalioxylon Schönfeld 1947 el tipo de porosidad, tipos de parénquima, el tipo de puntuaciones y el ancho de los radios.
Se establecieron comparaciones con 22 especies neógenas de Terminalioxylon distribuidas en Colombia, África, India y Argentina distinguidas por la presencia de porosidad difusa y radios uniseriados a biseriados (ver tabla Tabla 3 en Ramos et al. 2012 y tabla 4 en Kloster et al. 2017). No obstante, el juego de combinaciones del parénquima axial, las fibras, la presencia/ausencia de canales traumáticos y los elementos vasculares acercan este material a Terminalioxylon sp. Lemoigne 1978 del Terciario de Etiopía. Sin embargo, su descripción tiene interrogantes en cuanto a las fibras, canales traumáticos y el tipo de radios; razón por la cual, se considera necesario crear una nueva especie de Terminalioxylon: T. calchaquii para el Mioceno de la Formación San José ya que el material cuenta con los caracteres diagnósticos pertinentes.
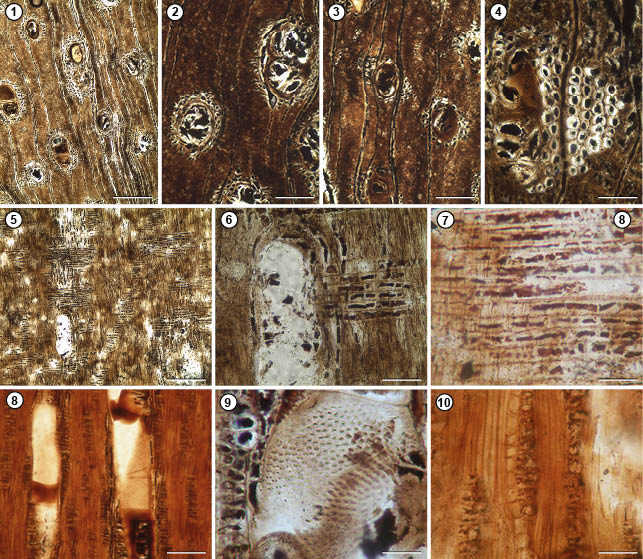
Figura 5. Terminalioxylon calchaquii nov. sp. 1. Lill PB: 045, 046, 047. 045. Corte transversal (1–4). 1. Anillos de crecimiento no marcados, porosidad difusa, tipo de vasos; en 2–4. Vasos con contenido; parénquima paratraqueal vasicéntrico a confluente con tendencia a aliforme. Fibras de contorno poligonal. 046. Corte longitudinal radial 5–7. Radios homocelulares con células procumbentes con cristales. 047. Corte longitudinal tangencial (8–10). 8. Placas de perforación simples. 9. Detalle de las punteaduras intervasculares. 10. Seriación de los radios; parénquima septado con cristales (flecha). Escalas: 1, 5= 150 μm; 2, 3,8= 100 μm; 4, 6, 10= 80 μm; 7, 9= 60 μm.
Tabla 1. Síntesis paleoecológica sobre la Formación San José (Mioceno medio).
|
Edad |
Mioceno medio (Gavriloff y Bossi, 1992; Bossi et al. 1997; Gavriloff, 1999; Kleinert y Strecker, 2001; Morton y Herbst, 2003) |
|
Asociación megaflorística |
Hojas de Malvaciphyllum, Myrciophyllum, etc (Anzótegui y Cristalli, 2000 y Anzótegui, 2002). Frutos: Asterocharpus, Baccharis, Cyperocarpus (Anzótegui y Herbst, 2004) |
|
Paleoambiente |
Unidad fluvio-lacustre con cuerpos de agua (lagos) y secciones palustres, rodeados de un bosque higrófilo y xerófilo (Anzótegui y Herbst, 2004) |
|
Paleoclima |
Estacionalidad climática sugerida por las estructuras sedimentarias presentes. Mayoría de leños con porosidad difusa pudiéndose interpretar esa disposición como una adaptación para optimizar el transporte en períodos secos |
|
Asociación de maderas |
Tiupuncoxylon angiona, Tabebuixoxylon eorigida, Terminalioxylon calchaquii |
|
Asociación microflorística |
Cordiapollenites, Euphortricolporites y Prosocolporites, Margocolporites, Acaciapollenites, Polyadopollenites (Mautino 2010 y Mautino y Anzótegui, 2014) |
consideraciones finales
La xiloflora de la Formación San José aporta evidencia significativa sobre las condiciones paleoambientales y paleoclimáticas del Grupo Santa María (Cuadro 1).
El índice de Vulnerabilidad (IV) para las especies descriptas fue de 8, 9,56 y 10. Estos valores sugieren que las especies poseen una buena capacidad de conducción de agua, con sensibilidad a condiciones de estrés hídrico, como sequías prolongadas o heladas intensas, debido a la mayor susceptibilidad de los vasos grandes a la cavitación. El Índice de Mesomorfía (IM) dieron valores de 1233,24, 4200 y 4080, lo que permite caracterizar a las especies como mesomórficas.
Las características anatómicas observadas como la porosidad difusa, las punteaduras intervasculares alternas y la mesomorfía de las especies descriptas en este trabajo indican eficiencia en el transporte de agua, lo que refuerza la hipótesis de un ambiente con variaciones hídricas estacionales y coincide con estudios previos sobre el entorno deposicional de esta unidad (García y Herbst, 1997; Morton y Herbst, 2003; Mautino, 2010; Mautino y Anzótegui, 2014; Robledo et al., 2017).
Finalmente, la diversidad taxonómica mega y microflorística registrada en la Formación San José la posiciona como una de las asociaciones más heterogéneas dentro del Grupo Santa María.
conflicto de intereses
Las autoras declaran no tener conflictos de intereses.
Agradecimientos
Las autoras desean expresar su sentido agradecimiento a la memoria de los Dres. Rafael Herbst y Alicia Lutz por la orientación y cariño que les brindaran. Hacen extensivo este reconocimiento a la Dra. Luisa Anzótegui que continúa distinguiéndose por el afecto que prodiga a los cultores de la Paleobotánica, y a los miembros de la Comunidad Amaicha del Valle por la gentileza y cordialidad de su atención. Asimismo, valoran el apoyo del PI2022 F024 de la SGCyT-UNNE para la realización de este trabajo. Las autoras quieren destacar la cuidadosa labor de los revisores que contribuyeron a mejorar la calidad de este manuscrito.
Contribuciones de los autores
Baez, J. S.: determinación sistemática, análisis, interpretación, ilustración, redacción. y compaginación
Crisafulli, A.: análisis, interpretación, redacción y compaginación
Martinez, R. M.: análisis, interpretación, redacción y compaginación
Todas las autoras leyeron y aprobaron el manuscrito final.
Anzótegui, L. M. (1997). Análisis preliminar de la paleoflora de la Formación San José (Neógeno) del Valle de Santa María (Tucumán), Argentina. Geociencias Resumen, 2, 238.
Anzótegui, L. M. (2001). Bulnesia (Zygophyllaceae) en la Formación San José (Mioceno Medio), provincia de Tucumán, Argentina. Boletín de la Sociedad Argentina de Botánica, 36, 136.
Anzótegui, L. M. (2002). Hojas de Myrtaceae en la Formación San José (Mioceno medio), provincia de Tucumán, Argentina. Ameghiniana, 39(3), 321–330.
Anzótegui, L. M., & Cristalli, P. (2000). Primer registro de hojas de Malvaceae en el Neógeno de Argentina y Brasil. Ameghiniana, 37(2), 169–180.
Anzótegui, L. M., & Ferraro, L. I. (2000). Líquenes folícolas en la Formación San José (Mioceno Medio), Tucumán, Argentina. Geociencias, 5, 256.
Anzótegui, L. M., & Herbst, R. (2004). Megaflora (hojas y frutos) de la Formación San José (Mioceno Medio) en Río Seco, departamento Santa María, provincia de Catamarca, Argentina. Ameghiniana, 41(3), 423–436.
Anzótegui, L. M., Mautino, L. R., Horn, M. Y., Garralla, S. S., Robledo, J. M., Nasif, N., ... & Georgieff, S. (2019). Paleovegetación del Mioceno tardío del noroeste de Argentina. Opera Lilloana, 52, 109–130.
Anzótegui, L. M., & Morton, L. S. (2000). Capítulos de Asteraceae en la flora de la Formación San José (Mioceno Medio), Tucumán, Argentina. En Reunión anual de Comunicaciones de la Asociación Paleontológica Argentina (Ameghiniana, 37(4)). Mar del Plata, Argentina.
Avise, J., & Johns, G. (1999). Proposal for a standardized temporal scheme of biological classification for extant species. Proceedings of the National Academy of Sciences of the United States of America, 96, 7358–7363.
Baez, J. S. (2022). Estudios xilotafoflorísticos del Neógeno de los Valles Calchaquíes, Argentina [Tesis doctoral inédita]. Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste.
Baez, J., Crisafulli, A., Martínez, R. M., & Mautino, L. (2019b). Avances en el conocimiento de la xilotafología neógena del noroeste argentino. En Reunión de Comunicaciones de la Asociación Paleontológica Argentina (83). La Plata, Argentina, 27–29 de noviembre.
Baez, J., Crisafulli, A., & Martínez, R. M. (2021). Novedades en la xilotafoflora de la Formación San José (Neógeno) de Tiu Punco, Tucumán, Argentina. En VI Simposio del Mioceno-Pleistoceno del Centro y Norte de Argentina (30 de noviembre–3 de diciembre). Buenos Aires, Argentina.
Benítez, M. H. (2022). Identificación de parámetros bioestratinómicos en niveles bioclásticos de la Formación San José (Mioceno). En Libro de Resúmenes de la Reunión de Comunicaciones de la Asociación Paleontológica Argentina 2022 (84). Publicación Electrónica de la Asociación Paleontológica Argentina, 23(1R), R1–R160.
Bossi, G. E., & Palma, R. (1982). Reconsideración de la estratigrafía del valle de Santa María, provincia de Catamarca, Argentina. En 5º Congreso Latinoamericano de Geología, Actas (Vol. 1, pp. 155–172). Buenos Aires, Argentina.
Bossi, G. E., Muruaga, C. M., Georgieff, S. M., Ahumada, A. L., Ibañez, L. M., & Vides, M. E. (1997). The Santa María Neogene Basin of the Pampean Ranges: An example of mixed tectonic evolution. En 1º Congreso Latinoamericano de Sedimentología (Vol. 1, pp. 97–104). Sociedad Venezolana de Geólogos, Caracas, Venezuela.
Brea, M., Zucol, A. F., & Patterer, N. (2010). Fossil woods from late Pleistocene sediments from El Palmar Formation, Uruguay Basin, Eastern Argentina. Review of Palaeobotany and Palynology, 163, 35–51.
Bromhead, A. (1840). Magazine of Natural History, n.s., 4, 333.
Brown, R. (1810). Prodromus florae Novae Hollandiae (Vol. 1, p. 260). London: Johnson.
Carlquist, S. (1977). Onagraceae: Additional species and concepts. Annals of the Missouri Botanical Garden, 64, 627–637.
Carlquist, S. (2001). Comparative wood anatomy: Systematic, ecological and evolutionary aspects of dicotyledon wood. Springer Series in Wood Science. Springer.
Collinson, M. E. (1986). Use of modern generic names for plant fossils. En R. E. Spicer & B. A. Thomas (Eds.), Systematic and taxonomic approaches in palaeobotany (Special Volume No. 31, pp. 91–104). The Systematic Association, Clarendon Press, Oxford.
Cozzo, D. (1964). Glosario de términos empleados en anatomía de maderas. Revista de la Facultad de Agronomía y Veterinaria, Universidad de Buenos Aires, 16, 33–79
Chase, M. W., Christenhusz, M. J. M., Fay, M. F., Byng, J. W., Judd, W. S., Soltis, D. E., & Stevens, P. F. (2016). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society, 181, 1–20.
Chattaway, M. (1932). Proposed standards for numerical values used in describing woods. Tropical Woods, 29, 20–28.
Ewers, F. W., Jacobsen, A. L., & López-Portillo, J. (2023). Carlquist’s indices for vulnerability and mesomorphy of wood: Are they relevant today? IAWA Journal, 44(3–4), 355–367.
Franco, M. J., & Brea, M. (2010). Microlobiusxylon paranaensis gen. et sp. nov. (Fabaceae, Mimosoideae) from the Pliocene-Pleistocene of Ituzaingó Formation, Paraná Basin, Argentina. Review of Palaeobotany and Palynology, 162, 123–132.
Franco, M. J., & Brea, M. (2013). Leños fósiles de leguminosas de la Formación Ituzaingó, Entre Ríos, Argentina: Implicancias paleoecológicas y paleobiogeográficas. Ameghiniana, 50(2), 167–191.
Galván, A. F., & Ruiz Huidobro, O. (1965). Geología del Valle de Santa María: Estratigrafía de las formaciones terciarias. Acta Geológica Lilloana, 7, 217–230.
García, A., & Herbst, R. (1997). Charophyta del Mioceno Superior de los valles de Santa María y de El Cajón, provincia de Catamarca, Argentina. Ameghiniana, 34(1), 61–68.
Georgieff, S. M., Ibañez, L. M., Vides, M. E., Anis, K. B., & Nieva, S. M. (2014). Paleógeno y Neógeno de Tucumán: Estratigrafía y paleoambientes sedimentarios. En Geología de Tucumán (pp. 106–123).
Gregory, M. (1994). Bibliography of systematic wood anatomy of dicotyledons. IAWA Journal, 1, 1–265.
Gregory, M., Poole, I., & Wheeler, E. (2009). Fossil dicot wood names: An annotated list with full bibliography. IAWA Journal, 6, 1–128.
Herbst, R. (2016). Paleobiogeografía del valle de Yocavil. Instituto Superior de Correlación Geológica (INSUGEO), 1–61.
Hass, H., & Rowe, N. P. (1999). Thin sections and wafering. En T. P. Jones & N. P. Rowe (Eds.), Fossil plants and spores: Modern techniques (pp. 76–81). Geological Society, London.
Herbst, R., Anzótegui, L. M., Esteban, G., Mautino, L. R., Morton, S., & Nassif, N. (2000). Síntesis paleontológica del Mioceno de los valles Calchaquíes, noroeste argentino. En F. Aceñolaza & R. Herbst (Eds.), El Neógeno de Argentina (INSUGEO, Serie Correlación Geológica, 14, pp. 263–288).
IAWA Committee. (1989). List of microscopic features for hardwood identification. IAWA Bulletin, 10, 219–332.
InsideWood. (2004–presente). InsideWood database. North Carolina State University. http://insidewood.lib.ncsu.edu/search
Jussieu, A. L. de. (1789). Genera plantarum, secundum ordines naturales disposita, juxta methodum in Horto Regio Parisiensi exaratam. Parisiis: Herissant et Barrois.
Kleinert, K., & Strecker, M. R. (2001). Climate change in response to orographic barrier uplift: Paleosol and stable isotope evidence from the Late Neogene Santa María Basin, northwestern Argentina. Geological Society of America Bulletin, 113(6), 728–742.
Lemoigne, Y. (1978). Flores tertiaires de la Haute Vallée de l’Omo (Éthiopie). Palaeontographica, 165B, 80–157.
Lindorf, H. (1994). Eco-anatomical wood features of species from a very dry tropical forest. IAWA Journal, 15(4), 361–376.
Lutz, A. I. (1987). Estudio anatómico de maderas terciarias del valle de Santa María (Catamarca-Tucumán), Argentina. FACENA, 7, 125–143.
Madel-Angeliewa, E., & Müller-Stoll, W. R. (1973). Kritische Studien über fossile Combretaceen-Hölzer: Über Hölzer von Typus Terminalioxylon G. Schönfeld mit einer Revision der bisher zu Evodioxylon Chiarugi gestellten Arten. Palaeontographica B, 142, 117–136.
Martínez, L. C. (2010). Prosopisinoxylon anciborae nov. gen. et sp. (Leguminosae, Mimosoideae) from the Late Miocene Chiquimil Formation (Santa María Group), Catamarca, Argentina. Review of Palaeobotany and Palynology, 158(3–4), 262–271.
Martínez, L. C. A. (2014). Fossil legume woods from the Late Miocene, Chiquimil Formation (Santa María Basin), Argentina. Review of Palaeobotany and Palynology, 201, 1–11.
Martínez, L. C. A., & Rodríguez-Brizuela, R. (2011). Gleditsioxylon riojana nov. sp. en el Neógeno de Argentina: Sus relaciones paleoambientales y biogeográficas. Geobios, 44(5), 461–472.
Mautino, L. R. (2007). Chlorophyta de los Valles Calchaquíes (Mioceno Medio y Superior), Argentina. Revista Española de Micropaleontología, 39(1–2), 81–102.
Mautino, L. R. (2010). Palinofloras de las Formaciones San José y Chiquimil (Mioceno Medio y Superior), noroeste de Argentina [Tesis doctoral inédita]. Universidad Nacional del Nordeste, Corrientes, Argentina.
Mautino, L. R., & Anzótegui, L. M. (2014). Novedades palinológicas de las Formaciones San José y Chiquimil (Mioceno medio y tardío), noroeste de Argentina. Revista del Museo Argentino de Ciencias Naturales, 16(2), 143–164.
Metcalfe, C. R., & Chalk, L. (1950). Anatomy of the Dicotyledons: Leaves, stem, and wood in relation to taxonomy, with notes on economic uses. Oxford: Clarendon Press.
Moglia, G., & Giménez, A. M. (1998). Rasgos anatómicos característicos del hidrosistema de las principales especies arbóreas de la Región Chaqueña Argentina. Forest Systems, 7(1), 53–71.
Morton, L. S., & Herbst, R. (2003). Moluscos dulceacuícolas de las Formaciones San José y Chiquimil (Mioceno) del Valle de Santa María (Catamarca y Tucumán), Argentina. Ameghiniana, 40(2), 205–216.
Morton, L. S., & Herbst, R. (2012). Etherioidea and Ampullarioidea (Mollusca) in the San José Formation (Middle Miocene), Santa Maria Valley, Salta Province, Argentina. Gaea: Journal of Geoscience, 8(2), 82–90.
Moya, E. V. (2018). Maderas de Leguminosae del Mioceno–Pleistoceno superior de la región noreste de la Argentina: Aspectos taxonómicos y su importancia como indicadores de las variaciones climáticas y biogeográficas [Tesis doctoral inédita]. Universidad Nacional de La Plata, Argentina.
Moya, E., & Brea, M. (2018). First Pleistocene record of fossil wood of Bignoniaceae in the Americas and a comparison with the extant Tabebuia alliance and Tecomeae. Botanical Journal of the Linnean Society, 187(2), 303–318.
Moyano, M. S. (2009). Análisis regional del emplazamiento, sedimentación y evolución sintectónica de los abanicos aluviales cuaternarios de las Sierras Pampeanas nordoccidentales (Catamarca y Tucumán) [Tesis doctoral inédita]. Facultad de Ciencias Naturales e IML, Universidad Nacional de Tucumán, Argentina.
Müller-Stoll, W. R., & Mädel, E. (1967). Die fossilen Leguminosen-Hölzer: Eine Revision der mit Leguminosen verglichenen fossilen Hölzer und Beschreibungen älterer und neuer Arten. Palaeontographica B, 121, 1–46.
Nasim, A., Babineau, M., Donovan Bailey, C., Hannah Banks, Barbosa, A., Barbosa Pinto, R., Boatwright, J., & Zimmerman, E. (2017). Legume Phylogeny Working Group: A new subfamily classification of the Leguminosae based on a taxonomic analysis. Phytotaxa, 300(2), 1–25.
Pace, M. R., & Angyalossy, V. (2013). Wood anatomy and evolution: A case study in the Bignoniaceae. International Journal of Plant Sciences, 174(7), 1014–1048.
Pujana, R. R. (2022). Fossil woods from Argentina (1884–2021). Revista del Museo Argentino de Ciencias Naturales (nueva serie), 24(2), 217–240.
Pujana, R. R., Martínez, L. C. A., & Brea, M. (2011). Maderas fósiles de Leguminosae en Sudamérica: Revisión del registro y afinidades sistemáticas. Revista del Museo Argentino de Ciencias Naturales, 13(2), 189–202.
POWO. (2024). Plants of the World Online. Royal Botanic Gardens, Kew. https://powo.science.kew.org/ (consultado el 23 de marzo de 2025).
Ramos, R. S., Brea, M., & Kröhling, D. (2012). Leños fósiles de la Formación El Palmar (Pleistoceno tardío) en el Parque Nacional El Palmar, Entre Ríos, Argentina. Ameghiniana, 49(4), 606–622.
Ramos, R. S., Brea, M., & Kröhling, D. (2017). Malvaceous wood from the Late Pleistocene El Palmar Formation of northeastern Argentina. Review of Palaeobotany and Palynology, 246, 232–241.
Robledo, J. M. (2017). Icnología de las asociaciones planta-insecto de paleofloras de las formaciones San José (Mioceno medio), Chiquimil y Palo Pintado (Mioceno tardío) de los Valles Calchaquíes [Tesis doctoral inédita]. Facultad de Ciencias Exactas, Naturales y Agrimensura, Universidad Nacional del Nordeste, Argentina.
Robledo, J. M., Pinheiro, E. R., Gnaedinger, S. C., & Wappler, T. (2018). Plant–insect interactions on dicots and ferns from the Miocene of Argentina. Palaios, 33(7), 338–352.
Ruiz, D. P., Pujana, R. R., & Brea, M. (2023). Paleocene fossil wood from Patagonia with storied rays and comments on the fossil record of this character. IAWA Journal, 45(1), 27–46.
Suguio, K., & Mussa, D. (1978). Madeiras fósseis dos aluviões antigos do Rio Tietê, São Paulo. Boletim do Instituto de Geociências da Universidade de São Paulo, 9, 25–45.
Schönfeld, G. (1947). Hölzer aus dem Tertiär von Kolumbien. Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, 475, 1–53.
WFO. (2020). World Flora Online. http://www.worldfloraonline.org (consultado el 25 de marzo de 2025).
Wheeler, E., & Baas, P. (1991). A survey of the fossil record for dicotyledonous wood and its significance for evolutionary and ecological wood anatomy. IAWA Bulletin, 12, 275–332.
Zuloaga, F. O., & Morrone, O. (Eds.). (1999). Catálogo de las plantas vasculares de la República Argentina. II. Angiospermae (Dicotyledoneae). Monographs in Systematic Botany from the Missouri Botanical Garden, 64, 1–1269.