Articulo original
Original article
36 (1) | Ene - Jun, 2026
Facultad de Ciencias Exactas y Naturales y Agrimensura (UNNE)
Open Access: https://revistas.unne.edu.ar/index.php/fce
E-mail: revistafacena@exa.unne.edu.ar
Morfología ovárica en neonatos de tres especies de lagartos ovíparos del nordeste de Argentina
Ovarian morphology in neonates of three oviparous lizard species from northeastern Argentina
Sandoval, María Teresa * ¹ , Arrieta, María Belén ¹, Delssin, Andrea Raquel ¹ & Alvarez, Blanca Beatriz
1 Laboratorio de Herpetología. Facultad de Ciencias Exactas y Naturales y Agrimensura. Universidad Nacional del Nordeste. Corrientes, Argentina.
* Autor de correspodencia: mtsandoval@exa.unne.edu.ar
Recibido/Received: 27 de noviembre, 2026 | Aceptado/Acepted: 01 de abril, 2026 | Publicado/Published: 08 de junio, 2026
Como citar este artículo: Sandoval, M. T., Arrieta, M. B., Delssin, A. R. & Alvarez, B. B. (2026). Morfología ovárica en neonatos de tres especies de lagartos ovíparos del nordeste de Argentina. Revista FACENA 36(1), 14-29. Doi: https://doi.org/10.30972/fac.3618981
Resumen: El desarrollo ovárico en reptiles constituye un proceso clave para comprender la biología reproductiva y los cambios ontogenéticos que ocurren antes de la maduración sexual. En este estudio se caracterizó la organización histológica del ovario de neonatos de tres especies de lagartos de la región nordeste de argentina: Salvator merianae, Tropidurus catalanensis y Cercosaura screibersii. Los resultados mostraron aspectos topográficos semejantes de los ovarios en las tres especies analizadas. A nivel histológico se registró una médula con lagunas bien desarrollada y una corteza con variaciones interespecíficas en el número y estadio de células germinales y folículos ováricos. S. merianae presentó escasas oogonias en la corteza, sugiriendo que la proliferación de la línea germinal y el inicio de la foliculogénesis ocurren en la etapa juvenil. En T. catalanensis y C. screibersii se observaron principalmente nidos de oocitos en fases iniciales de la meiosis, y folículos ováricos en estadios tempranos del desarrollo, lo que sería indicativo de que la foliculogénesis se inicia antes de la eclosión. T. catalanensis presentó, además, folículos previtelogénicos en crecimiento, característica no descripta para neonatos de otros reptiles. Los resultados sugieren variaciones interespecíficas en la morfogénesis ovárica y abre nuevas perspectivas de análisis del desarrollo y la maduración gonadal en escamados.
Palabras claves: Foliculogénesis; Histología; Ovario; Reptiles.
Abstract: Ovarian development in reptiles constitutes a key process for understanding reproductive biology and the ontogenetic changes that occur prior to sexual maturation. This study characterized the histological structure of the ovaries of neonates from three lizard species inhabiting the northeastern region of Argentina: Salvator merianae, Tropidurus catalanensis, and Cercosaura schrebersii. The results showed similar topographic features of the ovaries among the three species analysed. Histologically, a well-developed lacunar medulla and cortex were observed, but interspecific variation in the number and developmental stages of germ cells and follicles was noted. S. merianae presented few oogonia in the cortex, suggesting that germline proliferation and the onset of folliculogenesis occur during the juvenile stage. In T. catalanensis and C. schrebersii, nests of oocytes at early meiotic stages and follicles at early developmental stages were observed, suggesting that folliculogenesis begins before hatching. T. catalanensis also exhibited a growing previtellogenic follicle, a feature not previously described for neonates of other reptiles. These results show interspecific variation in ovarian morphogenesis and provide new approaches for the analysis of gonadal development and maturation in squamates.
Keywords: Folliculogenesis; Histology; Ovary; Reptiles.
Introducción
El desarrollo ovárico es un proceso fundamental en la biología reproductiva, caracterizado por procesos morfogenéticos y celulares que aseguran la formación de gametas funcionales y el mantenimiento de la fertilidad en las hembras. En los vertebrados, los ovarios se desarrollan a partir de un par de esbozos (crestas gonadales), derivados de un engrosamiento del epitelio celómico, localizados en la cara ventromedial del riñón mesonéfrico embrionario. Estas crestas son colonizadas por células germinales primordiales (CGP), de origen extragonadal, que se diferencian en oocitos los cuales se asocian a células foliculares para formar los folículos ováricos (Piprek et al., 2016).
Uno de los eventos clave en el desarrollo ovárico es la formación de nidos germinales que se originan por proliferación de oogonias en la corteza (Tingen y Woodruff , 2009). Estas agrupaciones son características de la etapa temprana de la morfogénesis y representan una condición filogenéticamente conservada tanto en vertebrados como en invertebrados (Saffman y Lasko, 1999). Radder et al., (2008) sugirieren que la organización espacial de las oogonias está ligada a la fecundidad de las especies y a la evolución de las estrategias reproductivas de los lagartos.
Desde un enfoque comparativo, se reconoce que, aunque la función básica del ovario es bastante conservada dentro de los amniotas, los mecanismos de diferenciación varían entre linajes, reflejando la evolución divergente de los programas morfogenéticos (DeFalco y Capel, 2009). Dentro del clado se reconoce una gran diversidad estructural y funcional, reflejo de adaptaciones evolutivas y ecológicas de cada grupo. En las aves y mamíferos, los ovarios son de tipo compacto con una médula formada por un estroma con abundantes vasos sanguíneos y una corteza donde maduran los folículos ováricos. En los peces teleósteos, anfibios y reptiles los ovarios son de tipo sacular y presentan un lumen lleno de linfa rodeado por una corteza de tejido conjuntivo vascularizado que contiene a las células germinales y folículos en desarrollo (Uribe Aranzábal, 2002).
Los escamados representan el grupo más diverso de saurópsidos diápsidos y uno de los menos estudiados en términos de morfogénesis gonadal. Se ha caracterizado la morfogénesis ovárica de Sceloporus undulatus (Austin, 1988), Calotes versicolor (Doddamani, 1994), Niveoscincus ocellatus (Neaves et al., 2006), Sceloporus aeneus (Antonio-Rubio et al., 2015), Tropidurus catalanensis (Delssin et al., 2019), Correlophus ciliatus, Eublepharis macularius, Lepidodactylus lugubris (Rams-Pociecha et al., 2023). Estos trabajos muestran que, sin bien los eventos del desarrollo ovárico son bastante conservados, existen diferencias interespecíficas en la morfología gonadal al final del desarrollo embrionario.
La estructura gonadal durante el periodo juvenil ha sido escasamente explorada en reptiles escamados, a pesar de que en esta etapa ocurren eventos clave de la maduración sexual e inicio de la actividad reproductiva. En el marco de la biología reproductiva de una especie, el conocimiento de la estructura ovárica durante el período juvenil resulta clave para comprender la dinámica ovárica e interpretar los eventos asociados a la madurez sexual de las hembras. El objetivo de este trabajo fue caracterizar la organización histológica del ovario de neonatos de Salvator merianae (Teiidae), Tropidurus catalanensis (Tropidurae) y Cercosaura screibersii (Gymnophthalmidae) y aportar datos de base que permitan profundizar el estudio de los eventos del desarrollo y maduración gonadal durante el periodo juvenil.
Materiales y métodos
Especies Seleccionadas
Para este trabajo se seleccionaron tres especies pertenecientes a diferentes familias y con características morfológicas, hábitos de vida y aspectos reproductivos diferentes, comunes en la región nordeste de Argentina.
Salvator merianae es un lagarto de gran tamaño perteneciente a la familia Teiidae. Los machos pueden alcanzar, en promedio, una longitud hocico-cloaca (LHC) de 376,6 mm y las hembras de 350,3 mm (Winck et al., 2011). En nuestro país, habita selvas húmedas tropicales y espacios abiertos de las provincias de Misiones, Corrientes, Chaco, Santa Fe, Buenos Aires, Córdoba, La Pampa y San Luis (Cei, 1993). Es ovípara con una fecundidad variable de 24 a 49 huevos por camada (Mercolli y Yanosky, 1995).
Tropidurus catalanensis pertenece a la familia Tropiduridade. Es común en ambientes abiertos y habitualmente se la observa sobre árboles, rocas o dunas arenosas con una amplia distribución en norte y centro de nuestro país (Cei, 1993; Teixeira y Giovanelli, 1999). La LHC promedio de los machos puede alcanzar los 101,2 mm y la de las hembras 89,7 mm. Es ovípara con una fecundidad variable de 4 a 9 huevos (Ortiz et al., 2014).
Cercosaura screibersii es una lagartija de pequeño tamaño de la familia Gymnophthalmidae que, en Argentina, se distribuye en Santiago del Estero, Corrientes, Misiones, Entre Ríos, Córdoba, San Luis y Buenos Aires. Habita generalmente debajo de rocas en pastizales o pajonales con vegetación arbustiva (Cei, 1993). Para las hembras se ha reportado una LHC promedio de 41,01 mm y para los machos de 35,59 mm (Schuck et al., 2021). Es una especie ovípara con una fecundidad fija de 2 huevos por cada camada (Diehl, 2007).
Material Biológico
Se realizaron cortes histológicos del ovario de tres neonatos (24 h post eclosión), uno de cada especie, provenientes de puestas incubadas en laboratorio. Los ejemplares fueron anestesiados y sacrificados con sobredosis de carticaina L-adrenalina siguiendo la guía de eutanasia animal de la American Veterinary Medical Association (AVMA, 2020). Dichos procedimientos contaron con el aval del Comité de Ética en Investigación de la FaCENA-UNNE (Resolución Nº 0756/18 C.D. FaCENA). A cada ejemplar se le midió la longitud hocico-cloaca (LHC) con calibre digital de 0,01mm de precisión. Mediante disección se aislaron los ovarios y se fijaron en solución de Bouin o formaldehído 10% tamponado. Las muestras se procesaron según la técnica histológica convencional consistente en: deshidratación en soluciones crecientes de alcohol etílico (70, 80 y 96%), alcohol butílico (100%), impregnación en alcohol butílico-parafina (50-50%) e inclusión en parafina (100%). Se realizaron secciones de 5-7 µm con micrótomo rotatorio tipo Spencer. Las muestras fueron coloreadas con Hematoxilina-Eosina (HE) y montadas con bálsamo sintético. Las observaciones y fotografías digitales se realizaron con lupa binocular Leica ES2 y microscopio óptico binocular Leica DM500. Las descripciones de la histología ovárica se realizaron con base en Uribe Aranzábal (2002); el reconocimiento de las células germinales y estadios (E.) de los folículos se realizó según Guraya (1989) y Uribe Aranzábal et al. (1995).
RESULTADOS
La LHC de los neonatos fue de 82,8 mm para Salvator merianae, 28,1 mm para Tropidurus catalanensis y 18,2 mm para Cercosaura screibersii.
A nivel macroscópico, en las tres especies los ovarios se observaron como estructuras ovoides y blanquecinas; su disposición fue simétrica en Salvator merianae (Figura 1a) y Tropidurus catalanensis (Figura 1b), y asimétrica en Cercosaura schrebersii (Figura 1c), donde el ovario derecho se ubicó más craneal que el izquierdo.
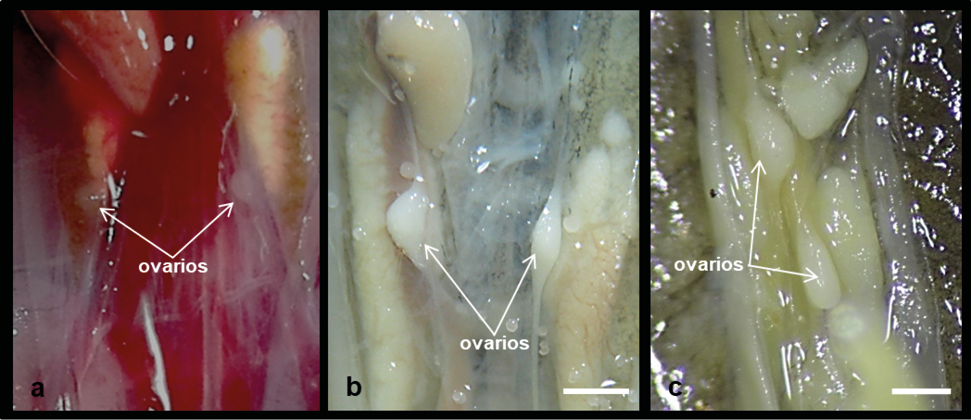
Figura 1. Morfología y topografía de los ovarios de neonatos. a) Salvator merianae, b) Tropidurus catalanensis y c) Cercosaura screibersii. Barra de escala: 1 mm.
A nivel histológico (Figura 2), el ovario presentó una médula con lagunas y una corteza bien desarrollada, delimitadas por una membrana basal, y rodeadas por un epitelio simple cúbico o escamoso. En las tres especies, la médula ovárica presentó abundante tejido conectivo, con vasos sanguíneos y lagunas medulares delimitadas por un epitelio simple cúbico en Salvator merianae (Figura 2d) y Cercosaura screibersii (Figura 2f) y un epitelio escamoso en Tropidurus catalanensis (Figura 2e).
En Salvator merianae la corteza ovárica presentó varias capas de células somáticas cúbicas o columnares, con núcleo ovoide o irregular. Dispersas en la corteza ovárica, se observaron escasas oogonias caracterizadas por su gran tamaño, citoplasma claro y núcleo voluminoso de contorno circular, con cromatina laxa (Figura 2g, j). Si bien se observaron algunas células germinales asociadas a células somáticas, no se registraron estructuras foliculares definidas.
En Tropidurus catalanensis la corteza ovárica presentó células somáticas columnares con núcleo ovoide alargado, numerosas oogonias formando nidos y oocitos primarios desnudos en fases iniciales de la meiosis caracterizados por la condensación de la cromatina (Figura 2h). Se observó, además, un folículo previtelogénico en E. III formado por el oocito y una capa folicular estratificada con células pequeñas e intermedias. El oocito se caracterizó por presentar un núcleo voluminoso, claro, con un nucléolo prominente y citoplasma homogéneo con algunas inclusiones lipídicas (Figura 2k).
En Cercosaura screibersii la corteza ovárica presentó escasas células somáticas y numerosos oocitos primarios desnudos en diplotene formando nidos germinales. Asimismo, se observaron algunos oocitos primarios rodeados parcialmente por células somáticas (prefoliculares) planas, correspondientes a folículos en E. 1 (Figura 2i, l).
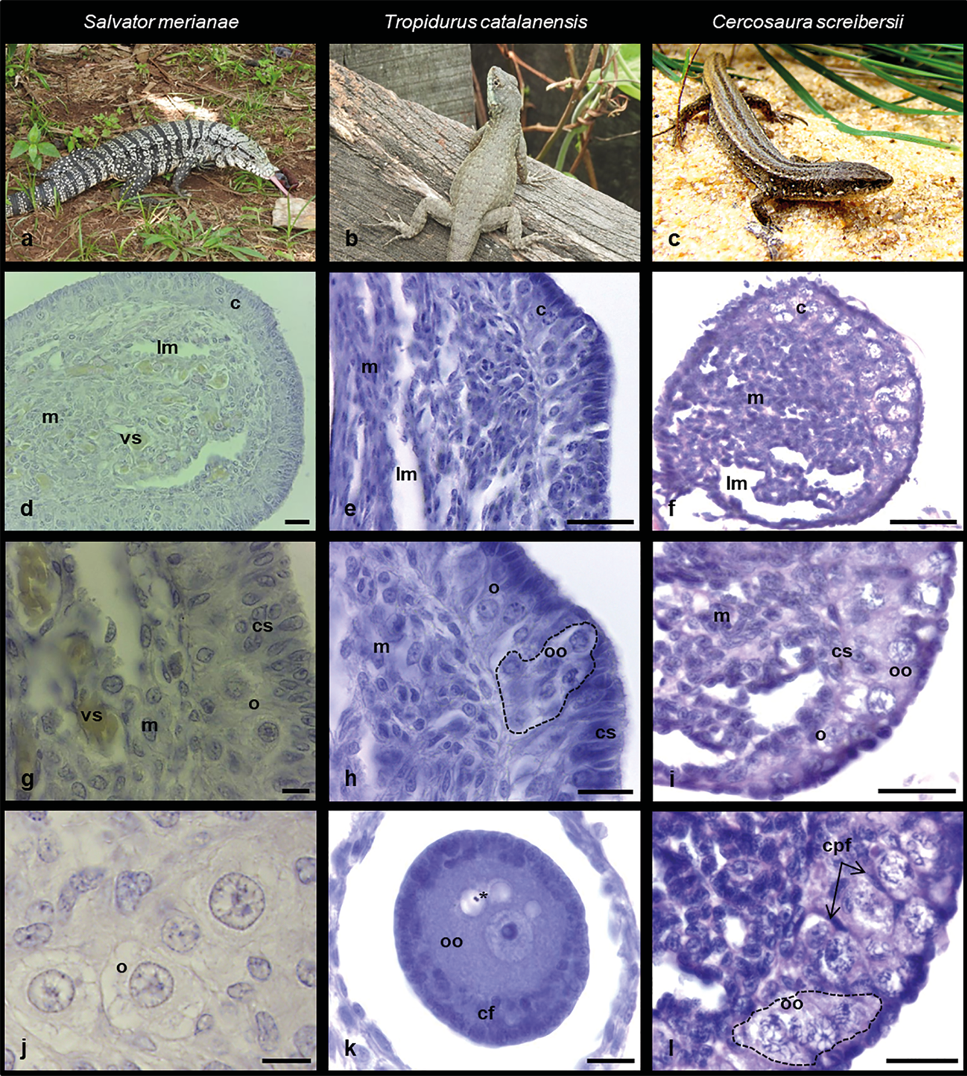
Figura 2. Histología ovárica de neonatos. Salvator merianae (columna izquierda), Tropidurus catalanensis (columna central) y Cercosaura screibersii (columna derecha). a-c: ejemplares adultos. d-f: aspecto del ovario. g-i: detalle de la corteza ovárica. j: detalle de oogonias. k: detalle de folículo ovárico. l: detalle de nidos de oocitos. Referencias: c: corteza, cpf: células prefoliculares, cs: células somáticas, lm: lagunas medulares, m: médula, o: oogonia, oo: oocito, (*): inclusiones lipídicas, línea punteada: nidos de oocitos. Barra de escala: 10 µm.
Discusión y conclusión
En los reptiles, la morfogénesis ovárica se inicia durante la organogénesis media del desarrollo embrionario siguiendo un patrón conservado en la mayoría de las especies (Ditewig y Yao, 2005). En el presente trabajo, la caracterización de la morfología ovárica de los neonatos permitió inferir los eventos que ocurren durante el periodo embrionario y post eclosión.
En Alligator mississippiensis (Moore et al.. 2008), Sceloporus aeneus (Antonio-Rubio et al. 2015), Sceloporus undulatus (Austin, 1988), Calotes versicolor (Doddamani, 1994), Niveoscincus ocellatus (Neaves et al., 2006), Correlophus ciliatus, Eublepharis macularius y Lepidodactylus lugubris (Rams-Pociecha et al., 2023) la formación de la cresta gonadal, la migración de las CGP, la diferenciación y proliferación de oogonias y el inicio de la meiosis ocurren durante el periodo embrionario, mientras que la formación de los folículos ováricos se produce luego de la eclosión. En Salvator merianae, la presencia de escasas oogonias en la corteza ovárica supone que la proliferación de la línea germinal, la meiosis y la folículogénesis se inician durante el periodo juvenil. La presencia de folículos en E. 1 en neonatos de Tropidurus catalanensis y Cercosaura screibersii podría indicar que la foliculogénesis se inicia antes de la eclosión en estas especies, lo que difiere de lo descripto para otras especies de lagartos. La observación de un folículo en E. III en Tropidurus catalanensis evidencia, además, el crecimiento folicular e inicio de la previtelogénesis, que incluyen la proliferación y diferenciación de las células foliculares y la inclusión de lípidos en el oocito, respectivamente. La presencia de folículos ováricos en desarrollo al finalizar la embriogénesis no ha sido previamente reportada para otros reptiles, aunque fue descripta para aves y mamíferos. En Coturnix coturnix (Rong et al., 2011) se describen folículos secundarios con 2 o 3 capas de células foliculares, al momento de la eclosión. En Sus scrofa domesticus, la oogénesis fetal incluye la formación de folículos primordiales y el crecimiento del oocito por depósito de inclusiones citoplasmáticas (Bielanska-Osuchowska, 2006). En mamíferos roedores, como Mus musculus y Rattus norvegicus, la foliculogénesis se inicia luego del nacimiento, en los primates Macaca mulatta y Homo sapiens los ovarios presentan folículos primordiales al final de la gestación y en rumiantes bóvidos como Bos taurus y Ovis orientalis se registran folículos preantrales, antrales y folículos atrésicos durante el periodo fetal (Hummitzsch et al., 2019).
La evidencia de lagunas medulares en los ovarios de los neonatos de las especies analizadas en este trabajo fue semejante a lo descripto para neonatos Niveoscincus ocellatus (Neaves et al., 2006) y embriones de Correlophus ciliatus, Eublepharis macularius y Lepidodactylus lugubris (Rams-Pociecha et al., 2023). En Sceloporus undulatus (Austin, 1988) y S. aeneus (Antonio-Rubio et al., 2015), al igual que en Chelonia mydas (Miller y Limpus, 2003), la médula permanece compacta hasta el final de la embriogénesis. Doddamani (1994) describe que en Calotes versicolor la cavitación medular ocurre a partir de los 10 días post eclosión.
La formación de lagunas medulares es una característica compartida entre reptiles y aves, resultado de la regresión de cordones sexuales primitivos en la región medular, y, al igual que la proliferación de la línea germinal y el inicio de la meiosis, está mediada por estrógenos (Pieau y Dorizzi, 2004; Ditewig y Yao, 2005). En mamíferos, el crecimiento y desarrollo folicular, previo al nacimiento, han sido asociados a una esteroidogénesis temprana (Pailhoux et al., 2001), mientras que en reptiles a un aporte hormonal materno (Smaga et al., 2023). El análisis de los perfiles de hormonas esteroides durante el desarrollo de los lagartos podría aportar datos que expliquen la ocurrencia de eventos característicos del periodo juvenil, en estadios embrionarios.
La variación en la temporalidad de los eventos del desarrollo ovárico (antes y luego de la eclosión) podría estar relacionada con diferencias en la biología reproductiva de los reptiles escamados. El grado de desarrollo al nacer, el tiempo de incubación intraovárica y estrategias energéticas de los embriones, condicionan los eventos de la morfogénesis ovárica. Demarco (1992) plantea que especies con mayor retención embrionaria o mayor grado de desarrollo al nacimiento tienden a mostrar etapas más avanzadas de foliculogénesis al momento de la eclosión. Los resultados de este trabajo sugieren la existencia de variaciones en la temporalidad y en la morfogénesis ovárica entre distintos linajes, lo que abre nuevas perspectivas para el análisis del desarrollo y la maduración gonadal en reptiles escamados. No obstante, dado que se analizó un único neonato por especie, estos hallazgos deben interpretarse como observaciones descriptivas de carácter preliminar. En este sentido, estudios con un mayor tamaño muestral serán necesarios para evaluar la variabilidad intraespecífica y corroborar los patrones aquí observados.
Conflicto de intereses
Las autoras declaran no tener conflictos de interés.
Agradecimientos
Un recuerdo y agradecimiento especial a Blanca Beatriz Álvarez quien nos brindó la posibilidad de iniciarnos en la actividad científica siendo un pilar fundamental en nuestra formación académica y profesional. Este trabajo fue financiado a partir de los proyectos 20F 002 y 24F 001 de la SGCyT-UNNE. Agradecemos al Dr. Víctor Hugo Zaracho por la fotografía del ejemplar adulto de Cercosaura screibersii y a los revisores de este manuscrito que han contribuido a mejorarlo sustancialmente.
En memoria de
Este trabajo se publica en memoria de la Lic. Blanca Beatriz Álvarez, quien contribuyó al desarrollo conceptual del estudio.
Contribuciones de los autores
Las autoras M. T. Sandoval, M. B. Arrieta y A. R. Delssin contribuyeron de manera equitativa a la obtención del material biológico, el procesamiento de muestras, el análisis y la descripción de las observaciones. Todas las autoras participaron en la redacción del manuscrito y en la preparación de las figuras.
Antonio-Rubio, N. R., Villagrán-Santa Cruz, M., Santos-Vázquez, A. y Moreno-Mendoza, N. (2015). Gonadal morphogenesis and sex differentiation in the oviparous lizard, Sceloporus aeneus (Squamata: Phrynosomatidae). Zoomorphology, 134(2), 279-289. https://doi.org/10.1007/s00435-015-0259-6
Austin, H. B. (1988). Differentiation and development of the reproductive system in the iguanid lizard, Sceloporus undulatus. General and comparative endocrinology, 72(3), 351-363. https://doi.org/10.1016/0016-6480(88)90157-8
AVMA. (2020). Guidelines for the euthanasia of animals: 2020 Edition. https://www.spandidos-publications.com/var/AVMA_euthanasia_guidelines_2020.pdf
Bielanska-Osuchowska, Z. (2006). Oogenesis in pig ovaries during the prenatal period: ultrastructure and morphometry. Reproductive Biology, 6(2), 161-193. https://www.pan.olsztyn.pl/repbiol/docs/pdfs/repbiol_vol6_num2_page161.pdf
Cei, J. M. (1993). Reptiles del noroeste, nordeste y este de la Argentina: Herpetofauna de las selvas subtropicales, Puna y Pampas. Museo Regionale di Scienze Naturali.Turino, Italia
DeFalco, T. y Capel, B. (2009). Gonad morphogenesis in vertebrates: divergent means to a convergent end. Annual Review of Cell and Developmental Biology, 25, 457-482. https://doi.org/10.1146/annurev.cellbio.042308.13350
Delssin, A. R., Sandoval, M. T., Ortiz, M. A. y Alvarez, B. B. (2019). Development and differentiation of the reproductive system of Tropidurus catalanensis (Squamata: Tropiduridae). Journal of Morphology, 280(2), 244-258. https://doi.org/10.1002/jmor.20940
Demarco, V. (1992). Embryonic development times and egg retention in four species of sceloporine lizards. Functional Ecology, 6(4),436-444. https://doi.org/10.2307/2389281
Diehl, L. S. (2007). Biologia reprodutiva de Cercosaura schreibersii (Wiegmann, 1834) (Sauria: Gymnophthalmidae) no sul do Brasil. (Tesis de Maestría, Pontifícia Universidade Católica do Rio Grande do Sul). https://repositorio.pucrs.br/dspace/bitstream/10923/5380/1/000392745-Texto%2BCompleto-0.pdf
Ditewig, A. C. y Yao, H. H. C. (2005). Organogenesis of the ovary: a comparative review on vertebrate ovary formation. Organogenesis, 2(2), 36–41. https://doi.org/10.4161/org.2.2.2491
Doddamani, L. S. (1994). Histoenzymological studies on embryonic and posthatching development of the ovary in the tropical oviparous lizard, Calotes versicolor. Journal of Morphology, 222(1), 1-10. https://doi.org/10.1002/jmor.1052220102
Guraya, S. S. (1989). Germinal beds, oogenesis and folliculogenesis. En Ovarian follicles in reptiles and birds (pp. 6-21). Berlin, Heidelberg: Springer Berlin Heidelberg.
Hummitzsch, K., Irving-Rodgers, H. F., Schwartz, J. y Rodgers, R. J. (2019). Development of the mammalian ovary and follicles. En The ovary (pp. ٧١-٨٢). Academic Press. https://doi.org/10.1016/B978-0-12-813209-8.00004-2
Mercolli, C. y Yanosky, A. A. (1995). Incubación artificial de huevos de iguana overa (“Tupinambis teguixim”) (Sauria: Teiidae). Archivos de Zootecnia, 44(168), 379-389.
Miller, J. D. y Limpus, C. J. (2003). Ontogeny of marine turtle gonads. En P. L. Lutz, J. A. Musick y J. Wyneken (Eds), The Biology of Sea Turtles, (Vol. II, pp 199-220).
Moore, B. C., Uribe Aranzábal, M. C., Boggs, A. S. y Guillette Jr, L. J. (2008). Developmental morphology of the neonatal alligator (Alligator mississippiensis) ovary. Journal of Morphology, 269(3), 302-312. https://doi.org/10.1002/jmor.10583
Neaves, L., Wapstra, E., Birch, D., Girling, J. E. y Joss, J. M. (2006). Embryonic gonadal and sexual organ development in a small viviparous skink, Niveoscincus ocellatus. Journal of Experimental Zoology Part A: Comparative Experimental Biology, 305(1), 74-82. https://doi.org/10.1002/jez.a.249
Ortiz, M. A., Boretto, J. M., Piantoni, C., Álvarez, B. B. y Ibargüengoytía, N. R. (2014). Reproductive biology of the Amazon Lava Lizard (Tropidurus torquatus) from the Wet Chaco of Corrientes (Argentina): congeneric comparisons of ecotypic and interspecific variations. Canadian Journal of Zoology, 92(7), 643-655. https://doi.org/10.1139/cjz-2014-0002
Pailhoux, E., Mandon-Pepin, B. y Cotinot, C. (2001). Mammalian gonadal differentiation: the pig model. Reproduction Supplement, 58, 65-80. https://www.researchgate.net/profile/Eric-Pailhoux/publication/11389644_Mammalian_gonadal_differentiation_The_pig_model/links/61af6453fb3b7258a0a3ca85/Mammalian-gonadal-differentiation-The-pig-model.pdf
Pieau, C. y Dorizzi, M. (2004). Oestrogens and temperature-dependent sex determination in reptiles: all is in the gonads. Journal of Endocrinology, 181(3), 367-377. https://doi.org/10.1677/joe.0.1810367
Piprek, R. P., Kloc, M. y Kubiak, J. Z. (2016). Early development of the gonads: origin and differentiation of the somatic cells of the genital ridges. En Molecular mechanisms of cell differentiation in gonad development (pp. 1-22). Springer International Publishing.
Pontelo, T. P., Miranda, J. R., Felix, M. A. R., Pereira, B. A., da Silva, W. E., Avelar, G. F., Queiroz Mariano, F. C. M., Corrêa Guimarães, G. y Zangeronimo, M. G. (2018). Histological characteristics of the gonads of pig fetuses and their relationship with fetal anatomical measurements. Research in Veterinary Science, 117, 28-36. https://doi.org/10.1016/j.rvsc.2017.11.005
Radder, R. S., Pizzatto, L. y Shine, R. (2008). Morphological correlates of life-history variation: is lizard clutch size related to the number of germinal beds in the ovary. Biological Journal of the Linnean Society, 94, 81-88. https://doi.org/10.1111/j.1095-8312.2008.00959.x
Rams-Pociecha I., Mizia P. C. y Piprek R. P. (2023). Histological analysis of gonadal ridge development and sex differentiation of gonads in three gecko species. Biology, 13(1), 7. https://doi.org/10.3390/biology13010007
Rong, C., Guobin, C., Yurong, Q., Bichun, L. y Guohong, C. (2011). The development of ovary in quail’s embryo. African Journal of Biotechnology, 10(4), 712-717. https://www.ajol.info/index.php/ajb/article/view/92383
Saffman, E. E. y Lasko, P. (1999). Germline development in vertebrates and invertebrates. Cellular and Molecular Life Sciences, 55(8), 1141-1163. https://link.springer.com/article/10.1007/s000180050363
Schuck, L. K., Neely, W. J., Farina, R. K., Oliveira, J. M. y Tozetti, A. M. (2021). Morphological trait variation between two populations of Cercosaura schreibersii in Southern Brazil: Insights on Habitat-Driven Adaptation. Herpetological Conservation and Biology, 16(2), 337-344. https://www.researchgate.net/profile/Laura-Schuck/publication/354248790_Morphological_trait_variation_between_two_populations_of_Cercosaura_schreibersii_in_southern_Brazil_insights_on_habitat-driven_adaptation/links/6682bb07f3b61c4e2ca11628/Morphological-trait-variation-between-two-populations-of-Cercosaura-schreibersii-in-southern-Brazil-insights-on-habitat-driven-adaptation.pdf
Smaga, C. R., Bock, S. L., Johnson, J. M. y Parrott, B. B. (2023). Sex determination and ovarian development in reptiles and amphibians: from genetic pathways to environmental influences. Sexual Development, 17(2-3), 99-119. https://doi.org/10.1159/000526009
Teixeira, R. L. y Giovanelli, M. (1999). Ecologia de Tropidurus torquatus (Sauria: Tropiduridae) da restinga de Guriri, São Mateus, ES. Revista Brasileira de Biologia, 59, 11-18. https://www.scielo.br/j/rbbio/a/WCbwtjs5yQvwvVR7HGyfZRk/?format=html&lang=pt
Tingen C., Kim A. y Woodruff T. K. (2009). The primordial pool of follicles and nest breakdown in mammalian ovaries. Molecular Human Reproduction. 15(12), 795-803. https://doi.org/10.1093/molehr/gap073
Uribe Aranzábal, M. D. C. (2002). Atlas de histología de vertebrados. Universidad Nacional Autónoma de México.
Uribe Aranzábal, M. D. C., Mendez Omana, M. E., Gonzalez Quintero, J. y Guillette Jr, L. J. (1995). Seasonal variation in ovarian histology of the viviparous lizard Sceloporus torquatus torquatus. Journal of Morphology, 226(1), 103-119. https://doi.org/10.1002/jmor.1052260107
Winck, G. R., Blanco, C. C., y Cechin, S. Z. (2011): Population ecology of Tupinambis merianae (Squamata, Teiidae): homerange, activity and space use. Animal Biology, 61, 493-510. https://doi.org/10.1163/157075511X597647