Articulo original
Original article
36 (1) | Ene - Jun, 2026
Facultad de Ciencias Exactas y Naturales y Agrimensura (UNNE)
Open Access: https://revistas.unne.edu.ar/index.php/fce
E-mail: revistafacena@exa.unne.edu.ar
Diversidad y variación estacional de anfibios y reptiles de la cuenca media del arroyo Saladillo, sur de Santa Fe, Argentina
Diversity and seasonal variation of amphibians and reptiles in the middle basin of the Saladillo stream, southern Santa Fe, Argentina
Alesio, Cristian Javier * ¹ , Paiz, Daniel Alejandro ¹ & Rimoldi, Pablo Guillermo ¹
1 Catedra de Biología y Ecología, Facultad de Ciencias Veterinarias, Universidad Nacional de Rosario (UNR) Argentina.
* Autor de correspodencia: cjalesio@gmail.com
Recibido/Received: 18 de diciembre, 2025 | Aceptado/Acepted: 08 de abril, 2026 | Publicado/Published: 09 de junio, 2026
Como citar este artículo: Alesio, C. J., Paiz, D. A. & Rimoldi, P. G. (2026). Diversidad y variación estacional de anfibios y reptiles de la cuenca media del arroyo Saladillo, sur de Santa Fe, Argentina. Revista FACENA 36(1), 30-62. Doi: https://doi.org/10.30972/fac.3618995
Resumen: En este estudio se analizó la diversidad, riqueza y abundancia de la herpetofauna en diferentes ambientes que componen el paisaje de la cuenca media del arroyo Saladillo, Santa Fe, Argentina; además de las variaciones estacionales entre ellos. La cuenca se dividió en matriz con alto nivel de antropización, representada por campos agrícolas intensivos; matriz de bajo nivel de antropización, compuesta por campos diversificados con rotación agrícola-ganadera; parches de vegetación leñosa y el corredor biológico, representado por el arroyo Saladillo. Las actividades de muestreo se realizaron durante 2021 y 2022, abarcando todas las estaciones del año. Los resultados obtenidos suman un total de 281 registros de individuos, distribuidos en 3 órdenes, 8 familias y 16 especies, con mayor abundancia en las estaciones de primavera-verano. Es fundamental resaltar la necesidad de continuar con estudios ecológicos básicos que incluyan la elaboración de listados de especies y el monitoreo de estos grupos, especialmente en regiones donde la antropización y el avance de la agricultura intensiva ponen en riesgo a la fauna silvestre. Además, al no existir trabajos previos para el área de estudio, se desconoce cuáles son las especies presentes, y si estas han sufrido variaciones en los últimos años. Este tipo de investigaciones son fundamentales para comprender la biodiversidad local existente y poner en valor los ecosistemas, quienes juegan un papel crucial en la supervivencia de estas especies.
Palabras claves: Herpetología; Biodiversidad; Ecosistemas antropizados; Sur de Santa Fe.
Abstract: In this study, we analyzed the diversity, richness and abundance of herpetofauna and their seasonal variations in different environments that make up the landscape of the middle basin of the Saladillo stream. The basin was divided into a matrix with a high level of anthropization, represented by intensive agricultural fields; low level of anthropization matrix, composed of diversified fields with agricultural-livestock rotation; patches of woody vegetation and the biological of Arroyo Saladillo. Sampling activities were carried out during 2021 and 2022, covering all seasons of the year. The results obtained add up to a total of 281 records of individuals, distributed in 3 orders, 8 families and 16 species, with a greater presence in the spring-summer seasons. It is essential to highlight the need to continue with basic ecological studies that include the preparation of lists of species and the monitoring of these groups, especially in regions where anthropization and the advance of intensive agriculture put wildlife at risk. Furthermore, since there is no previous work for the study area, it is unknown which species are present, and if these have undergone variations in recent years. This type of research is essential to understand the existing local biodiversity and value the ecosystems, which play a crucial role in the survival of these species.
Keywords: Herpetology; Biodiversity; Anthropized ecosystems; South of Santa Fe Province.
.
Introducción
Los cambios en el uso y la cobertura del suelo derivados de las actividades humanas son considerados uno de los principales impulsores de la pérdida de biodiversidad a nivel global (Volpedo y Reyes 2008; Uribe Botero 2015; Brondizio et al., 2019). La transformación de los paisajes naturales implica alteraciones rápidas en las coberturas vegetales, fragmentación de ambientes y deterioro de los hábitats, procesos que afectan tanto a la fauna como a la flora y modifican las propiedades físicas y químicas del suelo (Veloza 2020). Este fenómeno se expresa con particular intensidad en América Latina, región donde los ecosistemas naturales están experimentando tasas aceleradas de pérdida y modificación por causas antrópicas (Almond et al., 2022), con consecuencias negativas sobre numerosas poblaciones de vertebrados.
En la región pampeana argentina, la expansión del modelo agrícola industrial produjo una reconfiguración casi total del paisaje original. La sustitución de la vegetación nativa por cultivos exóticos y la intensificación de labores agrícolas incluyendo el uso sostenido de agroquímicos, el laboreo del suelo y la compactación progresiva han generado uno de los territorios más transformados del país (Solbrig, 1999; Vera et al., 2021; Biasatti y Rimoldi 2022). Este proceso también ha desencadenado una marcada simplificación ecológica, vinculada principalmente al predominio de monocultivos y a la reducción de la heterogeneidad estructural del ambiente (Biasatti et al., 2019; Truitt, 2019). La pérdida de complejidad del paisaje afecta no solo la composición de especies vegetales y animales, sino también el funcionamiento de los ecosistemas, limitando su resiliencia frente a disturbios actuales y futuros (Bilenca et al., 2012).
La herpetofauna, que integra grupos de anfibios y reptiles, cumple funciones ecológicas fundamentales y participa en procesos clave que articulan ambientes terrestres y acuáticos (Valencia-Aguilar et al., 2012; Williams et al., 2022; Rodríguez Parra, 2023). Estos organismos colaboran en el control de poblaciones de invertebrados, sostienen flujos de energía dentro de las redes tróficas y contribuyen a ciclos ecológicos como la dispersión de semillas, la movilidad de nutrientes y el mantenimiento de cuerpos de agua (Williams y Tettamanti, 2024). Al mismo tiempo, su marcada sensibilidad a los cambios en las condiciones ambientales los convierte en indicadores eficaces del estado de los ecosistemas (Attademo et al., 2005). En la región pampeana, estos grupos están expuestos a factores de presión asociados al modelo productivo dominante, a la degradación de los humedales y a incrementos de radiación ultravioleta, además de impactos vinculados a la urbanización y a la ganadería (Kacoliris, 2024).
Los reptiles, por su parte, enfrentan amenazas adicionales relacionadas al uso directo por parte del ser humano, como la captura con fines comerciales, el mascotismo, la persecución por miedo o desconocimiento y la extracción para obtención de cueros o venenos (Ubeda y Grigera 2003). Estas presiones, sumadas a la pérdida de hábitat, comprometen la conservación de numerosas especies locales.
La realización de estudios sistemáticos sobre la diversidad de herpetofauna resulta esencial para comprender cómo estos organismos responden a la transformación del paisaje y para proponer medidas que favorezcan su persistencia (Kacoliris et al., 2022). En paisajes agrícolas, donde la matriz productiva domina la estructura espacial, adquiere especial relevancia analizar el papel de los ambientes remanentes parches, corredores y unidades diversificadas en la provisión de hábitats, la conectividad ecológica y la reproducción de las especies (Cushman, 2006).
En la provincia de Santa Fe, si bien los relevamientos herpetológicos se han incrementado en las últimas décadas, persisten amplias zonas con escaso conocimiento científico, especialmente en el sector sur. La cuenca del Arroyo Saladillo constituye un ejemplo de esta situación: un paisaje fuertemente modificado donde la distribución, abundancia y diversidad de anfibios y reptiles todavía no han sido documentadas con detalle.
En este marco, el presente estudio se propone caracterizar el ensamble de anfibios y reptiles en los distintos componentes del paisaje de la cuenca media del Arroyo Saladillo, evaluando además su variación estacional. Para ello se estimaron la riqueza específica, la diversidad, la abundancia relativa y la composición de especies en cada unidad ambiental, con el fin de aportar información que contribuya a comprender la dinámica de estos grupos en un paisaje altamente antropizado y avanzar hacia estrategias de conservación basadas en evidencia.
MATERIALES Y MÉTODOS
Descripción del área de estudio
El estudio se desarrolló en la cuenca media del Arroyo Saladillo, ubicada en el sur de la Provincia de Santa Fe, Argentina, y comprendida entre los 32°55’–33°30’ S, 60°35’–61°55’ O (Fig. 1). La cuenca abarca aproximadamente 3.100 km² y forma parte de la región Pampeana, dentro del complejo Pampa Ondulada. El arroyo sigue una dirección general OSO–ENE y desemboca en el río Paraná. El clima es templado cálido y húmedo, con un promedio anual cercano a 960 mm de precipitaciones, concentradas en verano. La región presenta suelos predominantemente agrícolas, bien drenados en la mayor parte de la cuenca media, y sectores con drenaje imperfecto en los valles de inundación. La red hídrica incluye varios cursos secundarios, entre ellos los canales Arroyo Canal Candelaria, Canal Sanford – Arequito, Arroyo Pueblo Álvarez, Arroyo La Adela – La Esperanza, Canal Bombal; entre los más importantes.
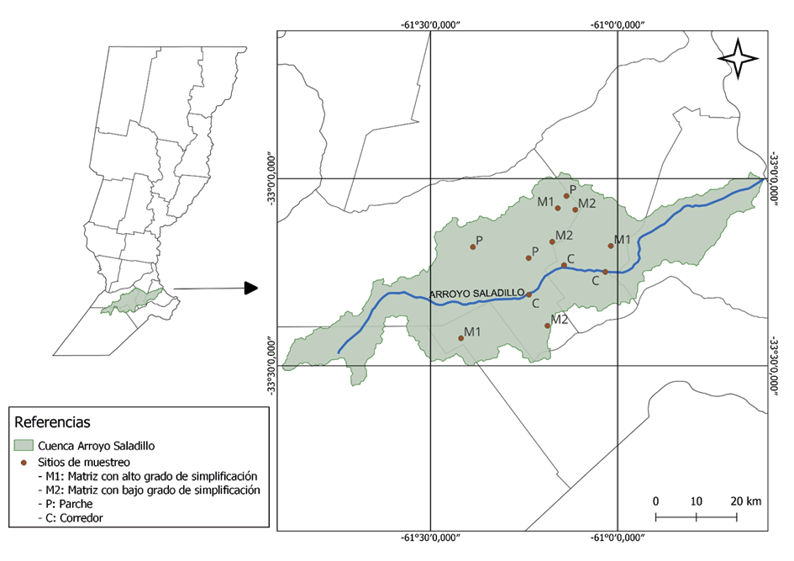
Figura 1. Delimitación del área de estudio en la cuenca media de Arroyo Saladillo. Cada punto representa un sitio de muestreo. Referencias: C (Corredor); M2 (Matriz con bajo grado de simplificación); M1 (Matriz con alto grado de simplificación); P (Parche).
Unidades del paisaje
Teniendo en cuenta que la heterogeneidad vegetal está directamente relacionada con el tamaño del área muestreada (Fox y Fox 2000, Lomolino y Perault 2001) y con el número de fragmentos de determinada cobertura o diversidad vegetal en la zona (Lomolino y Perault 2001) en este trabajo se establecieron 12 sitios de muestreo contemplando su rol de parche, matriz o corredor dentro del paisaje.
Los ambientes de muestreo fueron seleccionados en función de su representatividad dentro del mosaico de la cuenca media del Arroyo Saladillo y de la posibilidad de acceso con autorización de los propietarios. La asignación de los sitios a cada categoría se realizó mediante inspecciones de campo y verificación directa de las características estructurales de cada ambiente. Se definieron cuatro tipos de unidades del paisaje según su estructura y uso del suelo:
(1) Matriz agrícola simplificada (M1), dominada por monocultivos y alta intervención. Corresponde a una matriz agrícola altamente simplificada, caracterizada por la predominancia del doble cultivo trigo soja y la reducción casi total de las rotaciones agroganaderas tradicionales. En estos campos, la producción se basa mayormente en variedades de soja tolerantes a glifosato y en prácticas de siembra directa, lo que genera una estructura del paisaje homogénea, con escasa diversidad vegetal y elevada presión de manejo.
(2) Matriz diversificada (M2), con rotación agrícola-ganadera y mayor heterogeneidad de hábitats. En contraste, la matriz con bajo grado de simplificación reúne establecimientos con mayor heterogeneidad productiva y estructural. Estos predios combinan agricultura y ganadería, frecuentemente incorporan pasturas implantadas o naturales y pueden incluir producciones hortícolas o de pequeña escala (aves de granja, ovinos, equinos). La presencia de múltiples estratos vegetales, sumada a una flora más variada que en algunos casos incluye frutales, aromáticas y otras especies utilizadas en prácticas agroecológicas favorece la ocurrencia de artrópodos benéficos y genera ambientes más complejos que los campos estrictamente agrícolas.
(3) Parches de vegetación leñosa (P), asociados principalmente a montes remanentes o áreas peridomiciliarias. Estos ambientes lo constituyen los parches de vegetación leñosa, generalmente asociados a remanentes de antiguos cascos de estancias, montes implantados o construcciones abandonadas. Estos sectores presentan un claro predominio de especies arbóreas y arbustivas exóticas, como Broussonetia papyrifera, Eucalyptus spp., Melia azedarach, Ligustrum spp., Gleditsia triacanthos, Acacia melanoxylon u Olea europaea. Sin embargo, también pueden encontrarse ejemplares dispersos de especies nativas típicas de la región, incluyendo Erythrina crista-galli, Phytolacca dioica, Chorisia sp., Celtis tala, Enterolobium contortisiliquum, Tipuana tipu, Vachellia caven, Parkinsonia aculeata y Prosopis alba. Estos parches funcionan como pequeñas islas de refugio y aportan continuidad estructural dentro del paisaje agrícola.
(4) Corredor biológico (C) correspondiente al propio cauce y vegetación ribereña del Arroyo Saladillo. Este ambiente posee la mayor heterogeneidad dentro del sistema. A diferencia de la matriz circundante, dominada por cultivos extensivos, las márgenes del arroyo presentan una mayor complejidad vertical y horizontal, con una composición vegetal diversa y dinámica. Las comunidades ribereñas desempeñan un papel esencial en la provisión de microhábitats, la regulación ecológica y la conectividad funcional para múltiples grupos de fauna, lo que convierte a este corredor en un elemento clave para la conservación de los procesos ecológicos en la región.
Colecta de datos
El estudio se llevó a cabo durante los años 2021 y 2022 empleando una combinación de búsquedas visuales y métodos pasivos. Cada punto fue muestreado dos veces en cada estación del año, cubriendo así de manera sistemática los períodos estacionales relevantes para la actividad de anfibios y reptiles. Los relevamientos de anfibios se efectuaron mediante recorridas nocturnas entre las 19:00 y las 24:00 hs. utilizando lámparas frontales, mientras que los reptiles se registraron durante prospecciones matutinas realizadas aproximadamente entre las 09:00 y las 14:00 hs. En cada punto se dispusieron cuatro transectos lineales de 500 m, separados entre sí por 100 m, que fueron recorridos de manera sistemática. En todos los sitios se mantuvo un esfuerzo uniforme equivalente a una hora-hombre por muestreo, obteniendo los registros exclusivamente mediante observaciones directas, siguiendo las recomendaciones de Parris (1999), Angulo et al. (2006) y Gallina y López-González (2011). Totalizando dos horas-hombre de esfuerzo de muestreo por sitio-estación y grupo anfibios-reptiles.
Como complemento, se instalaron trampas cerco-pozo (pitfall traps), construidas con recipientes plásticos de 20 litros, de 28 cm de diámetro y 40 cm de profundidad, enterrados a ras del suelo. La base de cada recipiente fue perforada para facilitar el drenaje y se colocó un fragmento de goma espuma en el interior. En cada transecto se colocaron cuatro trampas en sitios considerados adecuados para la captura de herpetofauna, conforme a los lineamientos de Angulo et al. (2006). Cada batería de trampas estuvo constituido por cuatro recipientes dispuestos en forma de “Y”, conectados mediante tres barreras plásticas de aproximadamente 6 m de longitud, siguiendo el diseño propuesto por Pesci et al. (2018). Las trampas se mantuvieron activas una semana por estación y eran revisadas cada seis horas para minimizar la mortalidad de la fauna capturada. Al finalizar cada semana de muestreo, los recipientes se dejaban enterrados pero desactivados mediante tapas ajustadas para impedir la caída accidental de animales, siguiendo criterios similares a Perovic et al. (2008). En cada revisión se retiraron los ejemplares capturados, registrando la especie y el número de individuos antes de liberarlos en el mismo sitio. La abundancia relativa se expresó como número de individuos por trampa-día.
Consideraciones éticas: El proyecto fue aprobado por el Comité de Ética de la Facultad de Ciencias Veterinarias de la Universidad Nacional de Rosario (Código N° 18025/2020). Los ejemplares capturados fueron manipulados siguiendo los lineamientos éticos para el estudio de herpetofauna (ASIH, 2004). Todos los individuos fueron registrados y liberados en el mismo sitio de captura una vez finalizadas las tareas de identificación, priorizando su bienestar y minimizando el estrés durante los procedimientos.
Análisis estadísticos
Con el objetivo de caracterizar la diversidad en cada unidad del paisaje, se calcularon distintos parámetros derivados de la teoría clásica de diversidad. La riqueza específica (S) se definió como el número total de especies registradas por punto y por estación, siguiendo los criterios generales de estimación propuestos por Moreno (2001) y Magurran y McGill (2011). La equitatividad y la diversidad se obtuvieron mediante el índice de Shannon-Wiener, que permite evaluar la distribución relativa de las abundancias dentro de cada ensamblaje. Para complementar la interpretación ecológica de este índice, se transformaron los valores de H’ en números efectivos de especies mediante la expresión D = exp(H’), siguiendo a Jost (2006). Las abundancias utilizadas en estos cálculos corresponden a la integración de registros obtenidos mediante búsquedas visuales y trampas cerco-pozo, considerando ambos métodos como complementarios para la detección de especies.
La completitud del muestreo se evaluó mediante estimadores de diversidad alfa no paramétricos. Para cada unidad del paisaje se generaron curvas de acumulación de especies observadas y estimadas utilizando Bootstrap, Chao 2, ICE, Jackknife 2, así como los estimadores basados en especies únicas y duplicadas. Estos procedimientos no requieren supuestos distribucionales estrictos y permiten evaluar la suficiencia del esfuerzo de muestreo a partir de datos de presencia-ausencia.
La estructura de las comunidades se analizó mediante un escalamiento multidimensional no métrico (NMDS) aplicado a una matriz de abundancias transformadas mediante la distancia de Bray–Curtis. El análisis se realizó en dos dimensiones y se evaluó la estabilidad de la solución con 999 permutaciones, considerando el valor de stress como indicador de calidad de ajuste. Para contrastar diferencias en la composición de especies entre ambientes se empleó un PERMANOVA sobre la misma matriz de disimilitud, obteniendo el porcentaje de variación explicada (R²) y su significancia asociada. Estos procedimientos se implementaron en R versión 4.5.2 (R Core Team, 2025) utilizando los paquetes vegan y permute, mientras que las visualizaciones y la manipulación de datos se realizaron con ggplot2 y dplyr, respectivamente.
Las variables univariadas abundancia, riqueza y valores de Shannon-Wiener se compararon mediante un análisis de la varianza (ANOVA) bajo un modelo de efectos fijos con medidas repetidas, utilizando un nivel de significancia del 5%. Las estaciones del año fueron consideradas como factor intra-sujeto. Se verificaron los supuestos de normalidad mediante el test de Shapiro–Wilk y de homogeneidad de varianzas mediante el test de Levene. Esta aproximación se consideró adecuada para evaluar diferencias entre ambientes y estaciones en función del diseño balanceado del muestreo y de los objetivos comparativos del estudio. Cuando el ANOVA indicó diferencias estadísticamente significativas, se aplicó la prueba de comparaciones múltiples de Tukey para identificar qué grupos diferían entre sí, debido a su buen control del error tipo I en comparaciones por pares.
Adicionalmente, para las variables abundancia y riqueza se evaluó la interacción entre Ambiente y Estación mediante el análisis de la diferencia entre pares de estaciones (verano-invierno, otoño-invierno, primavera-invierno, etc.), utilizando ANOVA y pruebas de comparaciones múltiples de Tukey cuando correspondía. Esto permitió determinar si las variaciones estacionales diferían significativamente según el tipo de ambiente. Para la variable diversidad, al no encontrarse diferencias significativas en el factor Ambiente (p = 0,367), no se realizó el análisis de interacción. Estos análisis se realizaron con SPSS Statistics v27 (IBM, 2020).
RESULTADOS
Resultados generales
Los relevamientos realizados durante el 2021 y 2022 de anfibios y reptiles arrojaron un total de 281 registros de individuos, representados por 3 órdenes, 8 familias y 16 especies. Las cuales se encuentran mencionadas en la Tabla 1. En base al comportamiento de los estimadores de diversidad para cada uno de los ambientes parece improbable obtener un mayor número de especies que las colectadas, aunque se aumentara el esfuerzo de muestreo ya que las curvas de acumulación de especies se estabilizaron o tendieron a descender (Fig. 2)
Para cada una estas especies mencionadas se puede observar la presencia o ausencia, y la abundancia relativa para cada una de las unidades del paisaje que componen la cuenca media del arroyo Saladillo en la Tabla 2 y 3, estableciendo la diversidad alfa para cada ambiente.
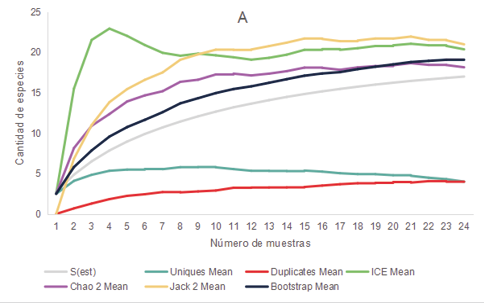
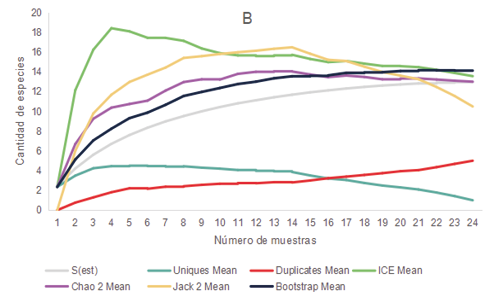
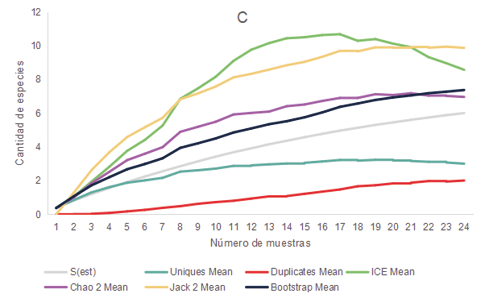
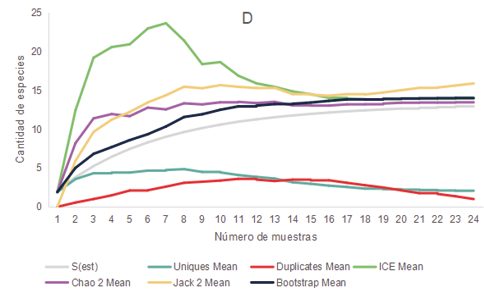
Figura 2. Curva de acumulación de especies de anfibios y reptiles (S obs) y curvas para los estimadores no paramétricos de riqueza Bootstrap, Chao 2, ICE, Jack 2, Uniques y Duplicates para los distintos ambientes presentes en los distintos ambientes de la cuenca media del arroyo Saladillo, sur de la provincia de Santa Fe, Argentina. A (Corredor) B (Parche) C (Campo agrícola) D (Campo diversificado).
Tabla 1. Especies de anfibios y reptiles relevadas en la cuenca media del arroyo Saladillo.
|
CLASE: AMPHIBIA |
|
|
Orden: Anura |
|
|
Familia: Microhylidae |
|
|
Elachistocleis bicolor (Guérin-Méneville, 1838) |
Sapito aceituna |
|
Familia: Hylidae |
|
|
Boana pulchella (Duméril & Bibron, 1841) |
Ranita de zarzal |
|
Familia: Leptodactylidae |
|
|
Leptodactylus gracilis (Duméril & Bibron, 1840) |
Rana rayada |
|
Leptodactylus latinasus (Jiménez de la Espada, 1875) |
Rana piadora |
|
Leptodactylus luctator (Hudson, 1892) |
Rana criolla |
|
Leptodactylus macrosternum (Miranda-Ribeiro, 1926) |
Rana criolla chaqueña |
|
Leptodactylus mystacinus (Burmeister, 1861) |
Rana de bigotes |
|
Physalaemus albonotatus (Steindachner, 1864) |
Rana maulladora |
|
Physalaemus biligonigerus (Cope, 1861) |
Ranita llorona |
|
Familia: Bufonidae |
|
|
Rhinella arenarum (Hensel, 1867) |
Sapo común |
|
Rhinella dorbignyi (Duméril & Bibron, 1841) |
Sapito cavador |
|
CLASE: REPTILIA |
|
|
Orden: Squamata |
|
|
Familia: Gymnophthalmidae |
|
|
Cercosaura schreibersii (Wiegmann, 1834) |
Lagartija negra |
|
Familia: Dipsadidae |
|
|
Erythrolamprus poecilogyrus (Wied-Neuwied, 1825) |
Culebra verde y negra |
|
Orden: Testudines |
|
|
Familia: Chelidae |
|
|
Phrynops hilarii (Duméril & Bibron, 1835) |
Tortuga de arroyo común |
|
Familia: Teiidae |
|
|
Salvator merianae (Duméril & Bibron, 1839) |
Lagarto overo |
|
Teius oculatus (d’Orbigny & Bibron, 1837) |
Lagartija verde común |
Tabla 2. Lista de anfibios y reptiles presentes en la cuenca media del arroyo Saladillo según registro de presencia por sitio de muestreo. Donde la X marca presencia.
|
NOMBRE CIENTÍFICO |
NOMBRE COMÚN |
SITIOS DE MUESTREO |
|||||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
||
|
Familia: Microhylidae |
|||||||||||||
|
Elachistocleis bicolor |
Sapito aceituna |
X |
X |
X |
X |
||||||||
|
Familia: Hylidae |
|||||||||||||
|
Boana pulchella |
Ranita de zarzal |
X |
X |
X |
X |
X |
|||||||
|
Familia: Leptodactylidae |
|||||||||||||
|
Leptodactylus gracilis |
Rana rayada |
X |
X |
X |
X |
X |
X |
||||||
|
Leptodactylus latinasus |
Rana piadora |
X |
X |
X |
X |
X |
X |
X |
X |
||||
|
Leptodactylus luctator |
Rana criolla |
X |
|||||||||||
|
Leptodactylus macrosternum |
Rana criolla chaqueña |
X |
X |
X |
X |
X |
|||||||
|
Leptodactylus mystacinus |
Rana de bigotes |
X |
X |
X |
X |
X |
X |
||||||
|
Physalaemus albonotatus |
Rana maulladora |
X |
X |
X |
X |
||||||||
|
Physalaemus biligonigerus |
Ranita llorona |
X |
X |
X |
X |
X |
|||||||
|
Familia: Bufonidae |
|||||||||||||
|
Rhinella arenarum |
Sapo común |
X |
X |
X |
X |
X |
X |
X |
X |
||||
|
Rhinella dorbignyi |
Sapito cavador |
X |
X |
X |
X |
X |
X |
X |
|||||
|
Familia: Gymnophthalmidae |
|||||||||||||
|
Cercosaura schreibersii |
Lagartija negra |
X |
X |
||||||||||
|
Familia: Colubridae |
|||||||||||||
|
Erythrolamprus poecilogyrus |
Culebra verde y negra |
X |
X |
X |
|||||||||
|
Familia: Chelidae |
|||||||||||||
|
Phrynops hilarii |
Tortuga de arroyo común |
X |
X |
X |
|||||||||
|
Familia: Teiidae |
|||||||||||||
|
Salvator merianae |
Lagarto overo |
X |
X |
X |
X |
X |
X |
X |
X |
X |
X |
X |
|
|
Teius oculatus |
Lagartija verde común |
X |
X |
X |
|||||||||
Tabla 3. Abundancia relativa de las distintas especies en los ambientes estudiados de la cuenca media del arroyo Saladillo, Santa Fe.
|
NOMBRE CIENTÍFICO |
TIPOS DE AMBIENTES |
|||
|
C |
P |
M1 |
M2 |
|
|
Familia: Microhylidae |
||||
|
Elachistocleis bicolor |
0,056 |
0,237 |
0 |
0,149 |
|
Familia: Hylidae |
||||
|
Boana pulchella |
0,135 |
0,022 |
0 |
0 |
|
Familia: Leptodactylidae |
||||
|
Leptodactylus gracilis |
0,022 |
0,075 |
0,083 |
0,184 |
|
Leptodactylus latinasus |
0,045 |
0,043 |
0,417 |
0,011 |
|
Leptodactylus luctator |
0,079 |
0 |
0 |
0 |
|
Leptodactylus macrosternum |
0,180 |
0,075 |
0 |
0,115 |
|
Leptodactylus mystacinus |
0,011 |
0,022 |
0,083 |
0,046 |
|
Physalaemus albonotatus |
0,101 |
0,290 |
0 |
0,103 |
|
Physalaemus biligonigerus |
0,034 |
0,065 |
0 |
0,092 |
|
Familia: Bufonidae |
||||
|
Rhinella arenarum |
0,236 |
0,032 |
0 |
0,011 |
|
Rhinella dorbignyi |
0,056 |
0,075 |
0 |
0,092 |
|
Familia: Gymnophthalmidae |
||||
|
Cercosaura schreibersii |
0,034 |
0 |
0 |
0,069 |
|
Familia: Dipsadidae |
||||
|
Erythrolamprus poecilogyrus |
0,034 |
0,043 |
0 |
0 |
|
Familia: Chelidae |
||||
|
Phrynops hilarii |
0,337 |
0 |
0 |
0 |
|
Familia: Teiidae |
||||
|
Salvator merianae |
0,180 |
0,140 |
0,417 |
0,092 |
|
Teius oculatus |
0,011 |
0,022 |
0 |
0,034 |
Comparación de las unidades del paisaje
El análisis NMDS mostró una clara segregación entre los cuatro sitios evaluados (C, M1, M2 y P), indicando diferencias marcadas en la composición de especies de anfibios y reptiles (stress = 0.145). Los sitios se agruparon de manera consistente en el espacio de ordenación, con el sitio C exhibiendo la mayor diferenciación respecto de M1, mientras que M2 y P presentaron composiciones más próximas entre sí. El PERMANOVA confirmó que el tipo de ambiente explicó una proporción significativa de la variación total observada (R² = 0.122, p = 0.001), evidenciando que cada sitio alberga una comunidad herpetológica distintiva (Fig. 3).
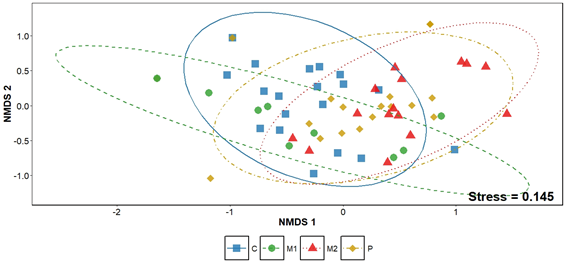
Figura 3. Análisis de ordenación no métrica multidimensional (NMDS) de la composición de especies de anfibios y reptiles en cuatro ambientes de la cuenca media del arroyo Saladillo, sur de la provincia de Santa Fe. Referencias: C (Corredor); M2 (Matriz con bajo grado de simplificación); M1 (Matriz con alto grado de simplificación); P (Pache).
En cuanto a la abundancia y la comparación entre los distintos ambientes se encontró que la abundancia de anfibios y reptiles observada en el ambiente M1 tuvo un promedio significativamente más bajo que el resto de los ambientes (x̅=1,04); y para el ambiente C un promedio significativamente más alto (x̅=5,75). Sin embargo, entre los ambientes M2, P y C la abundancia de anfibios y reptiles no presentó diferencias significativas.
Al igual que el parámetro riqueza, donde observamos que el ambiente M1 tuvo un promedio significativamente más bajo que el resto (x̅=0,79) y el ambiente C un promedio significativamente más alto (x̅=3,08). Sin embargo, entre los ambientes M2, P y C, la riqueza no presentó diferencias significativas.
En cuanto al índice de Shannon–Wiener, los valores obtenidos para el ambiente C fueron H’ 3,018, y para P, M1 y M2 fueron H’ 2,365, H’ 1,143 y H’ 2,281 respectivamente. Aunque, no resulta estadísticamente significativo, por lo tanto, se puede decir que los ambientes no difieren de manera significativa en cuanto a la diversidad observada de los anfibios y reptiles en cada uno.
La diversidad verdadera de anfibios y reptiles mostró diferencias marcadas entre las unidades del paisaje. Al expresar la diversidad como número efectivo de especies (exp(H’)), el Corredor (C) presentó el valor más alto (20.44 especies efectivas), seguido por el Parche (P) (10.64). En cambio, M2 exhibió una diversidad intermedia (9.78) y el ambiente agrícola M1 fue el menos diverso (3.14).
Patrones estacionales según unidades del paisaje
En cuanto a la variación de la estacionalidad para el parámetro abundancia la Fig. 4 (A) nos muestra que, al realizar las comparaciones de a pares de las estaciones estudiadas, se observa que hay diferencias significativas entre las estaciones, verano-invierno (p-valor = 0,000), otoño-invierno (p-valor = 0,019) e invierno-primavera (p-valor = 0,002), el resto de las comparaciones de a pares resultan no significativas.
Con el gráfico podemos confirmar los resultados de ANOVA y Tukey anteriores, en donde se encontró que entre el ambiente M1 se presenta diferencia significativa para la abundancia, con los ambientes M2, P y C. En estos últimos tres ambientes no se observan estas diferencias. También se observa que la estación de verano presenta diferencia significativa con invierno y este presenta a su vez diferencias significativas con otoño y primavera.
Además, se observa, por ejemplo, que el cambio entre verano e invierno en C, es mucho más marcado que en M1. El análisis de la interacción entre las estaciones invierno y verano, considerando diferentes ambientes, resultó significativo para el efecto ambiente (p-valor=0,000) en la disminución de la abundancia de anfibios y reptiles de verano a invierno. La disminución de abundancia de anfibios y reptiles de verano a invierno fue significativamente mayor en C y P, y menor en M1.
En cuanto al parámetro riqueza y la variación de la estacionalidad, al realizar las comparaciones por parejas de las estaciones estudiadas, se observa que hay diferencias significativas entre todas las estaciones, salvo entre otoño y primavera donde no se encuentra diferencia significativa (p-valor=0,347).
Al igual que con la variable abundancia, aquí también la Fig. 4 (B) confirma los resultados de ANOVA y Tukey anteriores para la variable riqueza, en donde se encontró que para el ambiente M1 se presentan diferencias significativas con el resto de los ambientes, y para los ambientes M2, P y C no se observan estas diferencias.
Además, se visualiza que el cambio entre verano y otoño en P es mucho más marcado que en M1. Y se puede ver que, durante la transición de verano a invierno, los ambientes P, C Y M2 mostraron una disminución significativamente mayor de la riqueza de anfibios y reptiles.
Cuando observamos las diferencias sobre la variación en la estacionalidad para el parámetro diversidad, se aprecia que solo el invierno difiere significativamente de verano y primavera. Para otoño no se encontró diferencia significativa con invierno, verano ni primavera (p-valor=0,104; p-valor=0,106 y p-valor=0,363; respectivamente). Verano y primavera tampoco difieren significativamente entre sí (p-valor=0,999) (Fig. 4 C).
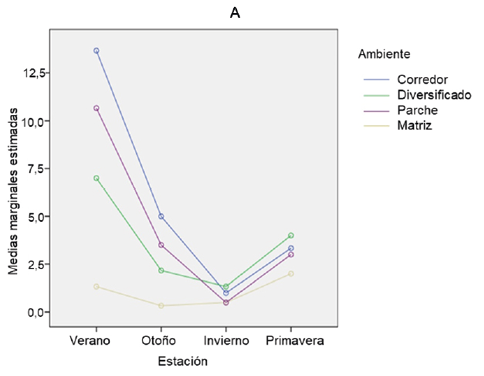
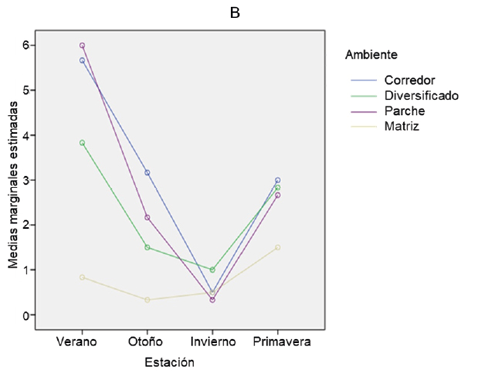
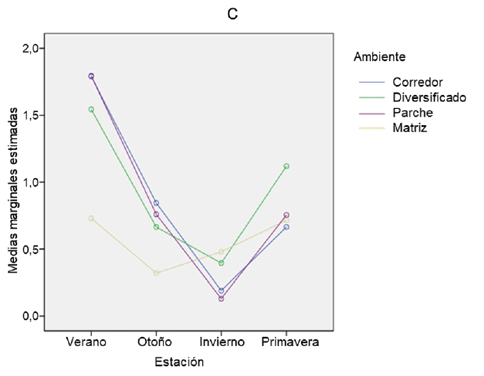
Figura 4. Comparación estacional de (a) abundancia, (b) riqueza y (c) diversidad de anfibios y reptiles en cuatro ambientes de la cuenca media del arroyo Saladillo, sur de la provincia de Santa Fe. Referencias: Ver (verano); Oto (otoño); Inv (invierno); Prim (primavera); C (Corredor); M2 (Matriz diversificada); M1 (Matriz agrícola simplificada); P (Parche de vegetación leñosa).
DISCUSIÓN
En el sur de la provincia de Santa Fe, la modificación del pastizal que caracterizaba a esta ecorregión ha desaparecido casi por completo, principalmente por el modelo agrícola-ganadero, que en las últimas décadas se ha profundizado hacia el modelo agrícola dominado por el monocultivo de soja (Glycine max) (Matteucci, 2012). En este ambiente (tierras de cultivos), el cual se presenta como matriz dominante del paisaje, es donde se registraron menos especies, pudiendo ser esto consecuencia de la profunda modificación del ecosistema que conlleva el modelo agrícola. Los campos diversificados (M2) junto a los parches (P) de vegetación leñosa albergaron especies de manera muy similar en términos de composición específica, mientras que el corredor biológico (C) presentó la mayor riqueza de especies. Estos resultados indican que los sectores con mayor cobertura vegetal y menor grado de intervención humana sostienen comunidades más diversas y estructuralmente complejas. Por el contrario, los ambientes más simplificados como M1 mantienen ensambles empobrecidos y dominados por un número reducido de especies generalistas o tolerantes a la perturbación.
La mayor diversidad registrada en el corredor biológico (C) puede atribuirse a una combinación de factores estructurales y funcionales propios de este ambiente. En primer lugar, la vegetación ribereña del arroyo Saladillo presenta una marcada heterogeneidad estructural, con una estratificación vertical y horizontal más compleja en comparación con las demás unidades del paisaje, lo que incrementa la disponibilidad y diversidad de microhábitats para anfibios y reptiles (Cushman, 2006). Esta complejidad incluye desde cuerpos de agua permanentes y semipermanentes, fundamentales para la reproducción de anuros, hasta refugios como troncos caídos, hojarasca y vegetación arbustiva densa, que ofrecen sitios de termorregulación y protección frente a depredadores (Peltzer et al., 2005). Asimismo, el corredor actúa como un eje de conectividad funcional dentro de una matriz agrícola altamente fragmentada, facilitando el movimiento de individuos entre parches y permitiendo el intercambio genético y el acceso a recursos estacionales (Baeza y Paruelo, 2020). La mayor humedad edáfica y atmosférica asociada a la presencia permanente del curso de agua también constituye un factor relevante, especialmente para anfibios, cuyas poblaciones dependen de condiciones de humedad para su actividad y supervivencia (Iriondo, 2007). En conjunto, estas características convierten al corredor ribereño en un refugio clave para la herpetofauna en un paisaje fuertemente antropizado, sustentando los mayores valores de riqueza, abundancia y diversidad registrados en este estudio.
En total, se registraron 11 especies de anfibios, siendo Leptodactylus luctator exclusiva del ambiente C. De las demás especies registradas, solo tres (L. gracilis, L. latinasus y L. mystacinus) estuvieron presentes en todos los ambientes relevados. Esto coincide con los resultados obtenidos por Suárez et al., (2016), quienes clasifican a estas especies, junto con Boana pulchella, como tolerantes a niveles de expansión e intensificación agrícola. Sin embargo, en este estudio, B. pulchella no se registró en los campos de cultivos agrícolas, aunque sí estuvo presente en los demás ambientes. Según López et al. (2015), esta especie puede adaptar su dieta en función de la disponibilidad de presas, lo que les otorga una ventaja adaptativa frente a las perturbaciones antropogénicas.
Por otro lado, Peltzer (2006), en coincidencia con los resultados obtenidos en esta investigación, encontró que L. latinasus fue una de las especies con mayor presencia en los campos de cultivo de soja, aunque la especie más frecuente fue P. albonotatus, registrada en todos los ambientes excepto en M1.
En cuanto a Elachistocleis bicolor, si bien las evaluaciones más recientes la clasifican como “Preocupación Menor” (LC) a nivel nacional (Vaira et al., 2012) y global (IUCN SSC Amphibian Specialist Group, 2023), su presencia en el área de estudio no fue homogénea entre los distintos ambientes. Esta especie fue detectada en el corredor biológico (C), en los parches de vegetación leñosa (P) y en la matriz diversificada (M2), pero estuvo ausente en la matriz agrícola simplificada (M1). Este patrón sugiere que, aunque se trata de una especie ampliamente distribuida y con tolerancia a cierto grado de modificación antrópica (Ghirardi y López, 2017), su persistencia en paisajes agrícolas podría estar condicionada por la presencia de hábitats con mayor heterogeneidad estructural y menor presión de agroquímicos.
La ausencia de E. bicolor en M1, sumada a su presencia en ambientes con mayor cobertura vegetal y diversidad de microhábitats, coincide con observaciones previas que indican que esta especie requiere suelos no compactados para su actividad fosorial y cuerpos de agua temporales para su reproducción (Lavilla, 2001; Ghirardi y López, 2017). En las matrices agrícolas simplificadas, la compactación del suelo asociada al tránsito de maquinaria agrícola y la reducción de sitios de reproducción por la desaparición de pequeños cuerpos de agua temporales podrían estar limitando su presencia. Por lo tanto, a pesar de su categoría de conservación favorable, la presencia de E. bicolor en paisajes altamente antropizados podría depender de la persistencia de ambientes con baja intervención, como los parches de vegetación y las matrices diversificadas, lo que refuerza la importancia de conservar estas unidades dentro del mosaico productivo.
En el grupo de reptiles se registraron cinco especies: Salvator merianae, Teius oculatus, Cercosaura schreibersii, Erythrolamprus poecilogyrus y Phrynops hilarii. La baja riqueza de reptiles en relación con los anfibios (11 especies) puede atribuirse en parte a diferencias en los requerimientos ecológicos y en la detectabilidad de cada grupo. Las especies registradas presentan distintos grados de tolerancia a la transformación del paisaje. Salvator merianae fue la especie de reptil con mayor distribución, presente en los cuatro tipos de ambientes, lo que coincide con su carácter generalista y su capacidad de utilizar hábitats modificados (Cabrera, 2015). En contraste, Phrynops hilarii fue exclusiva del corredor biológico, evidenciando su estrecha dependencia de cuerpos de agua permanentes, característica propia de las tortugas acuáticas (Cabrera, 2015). Teius oculatus y Cercosaura schreibersii, en cambio, se registraron únicamente en ambientes con mayor cobertura vegetal (P y M2), lo que sugiere requerimientos de hábitat más específicos vinculados a la disponibilidad de refugios y sustrato adecuado (Ubeda y Grigera, 2003). Cabe destacar que, previo a este estudio, Biasatti et al. (2016) solo habían mencionado para la cuenca del arroyo Saladillo la presencia de Salvator merianae, lo que subraya el carácter inédito del presente relevamiento y la importancia de continuar con estudios sistemáticos en la región.
Por otra parte, la ausencia de especies previamente citadas para la región, como Aspronema dorsivittatum, Ophiodes spp., Amphisbaena spp. o Philodryas patagoniensis (Cabrera, 2015), podría explicarse por la combinación de sus hábitos hipogeos o fosoriales y la metodología empleada, la cual priorizó registros visuales y trampas de caída superficiales (Angulo et al., 2006). Es probable que algunas de estas especies estén presentes en el área de estudio, pero no hayan sido detectadas debido a sus bajas tasas de encuentro o a la selectividad de los métodos de muestreo. Esta situación resalta la importancia de complementar los relevamientos con técnicas específicas para grupos crípticos (e.g., trampas de interceptación más profundas, búsqueda activa de refugios) y de continuar con estudios sistemáticos que permitan completar el inventario de la herpetofauna regional, especialmente en una zona con escasos antecedentes como la cuenca media del arroyo Saladillo.
Como se ha mencionado anteriormente, el ambiente M1 fue el menos utilizado por las especies relevadas. Este ambiente corresponde a campos dedicados exclusivamente a la agricultura intensiva, principalmente al monocultivo de soja. Este patrón coincide con lo reportado por Agostini et al. (2021), quienes encontraron menores valores de riqueza y abundancia de anuros en estanques asociados al cultivo de soja en comparación con otras áreas.
Diversos estudios han documentado que las prácticas asociadas a la agricultura intensiva pueden afectar a las comunidades de anfibios. Por ejemplo, se ha señalado que el uso de pesticidas puede impactar negativamente el desarrollo larval y la supervivencia de los adultos (Cuzziol Boccioni et al., 2024), así como modificar parámetros fisicoquímicos del agua como el pH, con potenciales efectos sobre el desarrollo y el comportamiento de los individuos (Peltzer, 2006). Asimismo, se ha observado que agroquímicos como el endosulfán pueden permanecer activos en medios acuáticos ácidos, intensificando su toxicidad (Lajmanovich et al., 2004), y que los fertilizantes pueden incrementar la concentración de nutrientes en cuerpos de agua, alterando su calidad y afectando el desarrollo larval (Bionda et al., 2012; Boone et al., 2004). En conjunto, estos factores, ampliamente documentados en la bibliografía, podrían estar contribuyendo a la menor riqueza y abundancia observada en los ambientes agrícolas más simplificados, aunque su incidencia específica en el área de estudio no fue evaluada directamente en este trabajo. Además, estudios previos sugieren que los efectos combinados de los agroquímicos pueden tener consecuencias más amplias sobre la estructura de las comunidades y el funcionamiento de los ecosistemas (Blaustein et al., 2010).
Por otra parte, la similitud de M2 con P, puede explicarse a partir de lo que menciona Altieri (1992) que sostiene que cuando hay diversificación productiva podría presentar diversidad de hábitats y mayor cantidad de recursos específicos para el desarrollo de la fauna, y heterogeneidad en el ambiente. En estos ecosistemas, además, la reducción en el uso de agroquímicos permite un mejor desarrollo de los anfibios (Lajmanovich et al., 2004, Peltzer et al., 2005). Aunque en este tipo de establecimientos se debe considerar que, en el caso de la ganadería, el sobrepastoreo puede traer consecuencias a los ecosistemas que afecta directamente a las poblaciones de anfibios (Agostini et al., 2021).
En cuanto a la estacionalidad y a la variación en los tres parámetros estudiados, se puede observar que los meses de septiembre a abril son los que han presentado los mayores registros. Esto se puede vincular a que estos meses coinciden con la estación reproductiva de las especies (Raffo, 2023), además, de la actividad trófica, lo que hace más fáciles de observar y escuchar (Peltzer, 2006). También debe considerarse que este es el período con la mayor cantidad de lluvias en la región, lo cual resulta fundamental para el éxito reproductivo de los anfibios, ya que el agua es un factor clave en su ciclo de vida (Iriondo, 2007).
Teniendo en cuenta esto, debemos considerar que tanto la riqueza como abundancia de anfibios encontrados podría ser mayor que la descripta, teniendo en cuenta que, durante los años de relevamientos, se registraron precipitaciones por debajo de la media, con valores anuales de 711 mm para el 2021 y 408 mm para el 2022 (registros obtenidos de la Estación Agrometeorológica de la Facultad de Ciencias Agrarias- UNR, Zavalla, Santa Fe). Estos valores muestran anomalías negativas de precipitación, donde la media esperada es de 1000 mm (Collins, 2022). Estas condiciones se condicen con los impactos esperados de “La Niña”, la fase fría del fenómeno océanico-atmosférico conocido como “El Niño- Oscilación del Sur”, que en la región de estudio está asociado a la falta de precipitaciones, especialmente en el semestre cálido (octubre a marzo) (Gastaudo et al., 2022).
La disponibilidad de agua es un factor determinante para el éxito reproductivo de los anfibios, ya que condiciona la existencia de cuerpos de agua con hidroperíodos suficientes para completar el desarrollo larval (Walls et al., 2013). En condiciones de déficit hídrico prolongado, las poblaciones pueden experimentar múltiples años consecutivos de fracaso reproductivo, con impactos negativos en la fecundidad y la supervivencia de los adultos (Walls et al., 2013). Estudios en ecosistemas de humedales efímeros han documentado que las sequías extremas reducen drásticamente la ocupación de sitios de reproducción y limitan el reclutamiento de nuevas cohortes (Awkerman y Greenberg, 2022). Asimismo, la disminución de la humedad ambiental afecta la actividad y detectabilidad de los individuos adultos, que dependen de microhábitats húmedos para su refugio y alimentación (Halstead et al., 2023). Si bien algunas especies pueden mostrar resiliencia a la variabilidad interanual, el déficit hídrico sostenido reduce la actividad de los adultos y compromete el éxito reproductivo (Halstead et al., 2023). Por lo tanto, es probable que las condiciones de sequía durante los años de muestreo hayan limitado tanto las oportunidades reproductivas como la detectabilidad de las especies, sugiriendo que los valores de riqueza y abundancia aquí reportados podrían ser subestimaciones de las comunidades realmente presentes en la cuenca en condiciones climáticas promedio.
Los resultados obtenidos en este estudio aportan evidencia cuantitativa que permite orientar estrategias de conservación en el sur de Santa Fe. Coincidiendo con Ubeda y Grigera (2003), quienes señalan que la vulnerabilidad de las especies de anfibios y reptiles está asociada a la destrucción del hábitat, nuestros resultados muestran que la matriz agrícola simplificada (M1) presentó los valores más bajos de riqueza (0,79 especies en promedio) y abundancia (1,04 individuos por muestreo), reflejando la marcada homogeneización del paisaje y la reducción de hábitats disponibles para la herpetofauna (Baeza y Paruelo, 2020). En contraste, el corredor biológico (C) sostuvo la mayor riqueza (3,08 especies en promedio) y abundancia (5,75 individuos), albergando especies exclusivas como Leptodactylus luctator y Phrynops hilarii, que no fueron registradas en otras unidades del paisaje. Por su parte, los parches de vegetación leñosa (P) y la matriz diversificada (M2) presentaron valores intermedios de riqueza y abundancia, y compartieron una composición de especies similar, lo que sugiere que estas unidades funcionan como hábitats complementarios dentro del paisaje productivo.
A partir de estos resultados, se destacan las siguientes prioridades de conservación:
Proteger y restaurar el corredor ribereño del arroyo Saladillo, dado que concentra la mayor diversidad y alberga especies con requerimientos ecológicos más específicos, no presentes en los ambientes agrícolas circundantes. La expansión de los monocultivos agrícolas produce inestabilidad y una disminución en la diversidad de hábitat (Altieri, 1994), mientras que la intensificación agrícola ha traído como consecuencia la disminución de parches y de conectividad entre los mismos (Baeza y Paruelo, 2020).
Mantener y potenciar los parches de vegetación leñosa (P) y las matrices diversificadas (M2), ya que estos ambientes, al presentar valores de riqueza y abundancia comparables entre sí, constituyen refugios clave en una matriz dominada por monocultivos. Una estrategia para disminuir el impacto en este tipo de ecosistemas es mantener la vegetación natural, ya que constituyen elementos del paisaje de gran importancia para la fauna, respecto de la oferta de refugio, conectividad estructural y funcional (Peltzer et al., 2005).
Revisar las prácticas de manejo en las matrices agrícolas simplificadas (M1). Si bien los mecanismos causales no fueron evaluados directamente en este estudio, la baja riqueza y abundancia registradas en estos ambientes, sumado a los antecedentes bibliográficos sobre los efectos de los agroquímicos en anfibios (Cuzziol Boccioni et al., 2024; Lajmanovich et al., 2004), sugieren la necesidad de implementar prácticas más sustentables que reduzcan el impacto sobre las comunidades silvestres. La creación de ambientes acuáticos en áreas agrícolas colaboraría con la preservación de los anuros (Peltzer et al., 2005), y conservando la vegetación en estos también se puede mitigar el impacto y producir una amortiguación de las consecuencias de la agricultura en la región (Peltzer, 2006).
En conjunto, estos hallazgos refuerzan la importancia de mantener la heterogeneidad del paisaje en agroecosistemas pampeanos, ya que la persistencia de la herpetofauna depende tanto de la preservación de los corredores biológicos y parches de vegetación como de la adopción de prácticas productivas que mitiguen los efectos negativos de la agricultura intensiva (Peltzer et al., 2005; Cushman, 2006). La pérdida de especies trae consecuencias ecológicas en los ecosistemas, ya que los anfibios cumplen un rol importante dentro de la cadena alimenticia, como controladores de insectos y como fuente de alimentos para otros (Cespedes et al., 2008).
Por último, es significativo resaltar la importancia de continuar con estudios ecológicos básicos de armado de listas de especies y monitoreo de estas como afirma Raffo (2023), sobre todo en regiones específicas, donde no se han realizado aún muestreos detallados de especies presentes en estos ambientes. Esto servirá además para conocer la biodiversidad presente y poner en valor a los ecosistemas que están cumpliendo un rol clave para la supervivencia de estas especies.
conclusiones
El presente estudio constituye el primer relevamiento sistemático de la herpetofauna en la cuenca media del arroyo Saladillo y evidencia que la transformación agrícola ejerce sobre estos grupos. La matriz agrícola simplificada (M1) fue el ambiente con menor riqueza, abundancia y diversidad, reflejando la reducción de hábitats, la homogeneización del paisaje. En contraste, el corredor biológico y los parches de vegetación leñosa sostuvieron las comunidades más diversas, destacando la importancia de estos ambientes como refugios y fuentes de conectividad ecológica dentro de un territorio altamente antropizado.
Los patrones estacionales mostraron que la mayor actividad y detectabilidad se concentran entre primavera y otoño, aunque la diversidad registrada podría estar subestimada debido al déficit hídrico asociado a eventos La Niña durante los años de estudio. La presencia de especies tolerantes a la perturbación en todos los ambientes y la limitada detección de reptiles reflejan tanto efectos del uso del suelo como limitaciones metodológicas.
En conjunto, los resultados muestran que es fundamental mantener los parches de vegetación y los corredores biológicos que todavía persisten en la zona, ya que funcionan como los principales refugios para la herpetofauna. También queda claro que sería necesario revisar algunas prácticas agrícolas, especialmente las relacionadas con el uso de agroquímicos, dado su impacto sobre estos grupos. Por último, este trabajo evidencia que aún falta información básica sobre la herpetofauna del sur de Santa Fe, por lo que continuar con estudios locales será clave para orientar acciones de manejo y conservación en un paisaje que ha cambiado de manera profunda.
Conflicto de intereses
Los autores declaran no tener conflicto de intereses.
Contribuciones de los autores
Cristian Javier Alesio: Conceptualización, metodología, investigación, análisis, escritura, revisión y edición.
Daniel Alejandro Paiz: Conceptualización, investigación y escritura.
Pablo Guillermo Rimoldi: Conceptualización, investigación, análisis, curaduría de datos y escritura.
Agostini, G., Deutsch, C. y Bilenca, D. N. (2021). Differential responses of anuran assemblages to land use in agroecosystems of central Argentina. Agriculture, Ecosystems & Environment 311: 107323. https://doi.org/10.1016/j.agee.2021.107323
Almond, R. E. A., Grooten, M., Juffe Bignoli, D. y Petersen, T. (Eds.). (2022). Informe Planeta Vivo 2022: Hacia una sociedad con la naturaleza en positivo. World Wide Fund for Nature.
Altieri, M. A. (1992). Biodiversidad, agroecología y manejo de plagas. Valparaíso, Chile: Ediciones CETAL.
Altieri, M. A. (1994). Biodiversity and pest management in agroecosystems. New York: Haworth Press.
American Society of Ichthyologists and Herpetologists (ASIH). (2004). Guidelines for Use of Live Amphibians and Reptiles in Field and Laboratory Research.
Angulo, A., Rueda-Almonacid, J. y Marca, E. (2006). Técnicas de inventario y monitoreo para los anfibios de la región tropical andina. Conservación Internacional. Serie Manuales de Campo Nº 2. Bogotá: Panamericana Formas e Impresos S.A.
Attademo, A., Lajmanovich, R., Peltzer, P. y Cejas W. (2005). Amphibians occurring in soybean and implications for biological control in Argentina. Agriculture, Ecosystems & Environment 106: 389-394.
Awkerman, J. A. y Greenberg, C. H. (2022). Projected climate and hydroregime variability constrain ephemeral wetland-dependent amphibian populations in simulations of southern toads. Ecologies 3(2): 235-248. https://doi.org/10.3390/ecologies3020018
Baeza, S. y Paruelo, J. M. (2020). Land use/land cover change (2000-2014) in the Rio de la Plata grasslands: an analysis based on MODIS NDVI time series. Remote Sensing 12(3): 381.
Biasatti, N.R., Rimoldi P.G. y Cabrera L.N. (2019). Desafíos de la conservación biológica en el Espinal y La Pampa Húmeda santafesina. Los mamíferos en el Sur de Santa Fe. Santa Fe (Argentina): Imprenta oficial de la provincia de Santa Fe.
Biasatti, N.R. y Rimoldi, P.G. (2022). Paradojas de la conservación de biodiversidad en los agroecosistemas pampeanos: la fragmentación inversa. Studies in Environmental and Animal Sciences 3(3): 754-761.
Biasatti, N.R., Rozzatti, J.C., Fandiño, B., Pautaso A., Mosso, E., Marteleur., Algarañaz, N., Giraudo, A., Chiarulli, C., Romano, M., Ramirez Llorens, P. y Vallejos, L. (2016). Las ecoregiones, su conservación y las áreas naturales protegidas de la provincia de Santa Fe. Santa Fe (Argentina): Imprenta oficial de la provincia de Santa Fe.
Bilenca, D., Codesido, M., González Fischer, C., Pérez Carusi, L., Zufiaurre, E. y Abba, A. (2012). Impactos de la transformación agropecuaria sobre la biodiversidad en la provincia de Buenos Aires. Revista del Museo Argentino de Ciencias Naturales 14(2): 189-198.
Bionda, C., Gari, N., Luque, E., Salas, N., Lajmanovich, R. y Martino, A. (2012). Ecología trófica en larvas de Rhinella arenarum (Anura: Bufonidae) en agroecosistemas y sus posibles implicaciones para la conservación. Revista de Biología Tropical 60(2): 771-779.
Blaustein, A. R., Walls, S. C., Bancroft, B.A., Lawler, J. J., Searle, C. L. y Gervasi, S. S. (2010). Direct and indirect effects of climate change on amphibian populations. Diversity 2(2): 281-313.
Boone, M. D, Semlitsch, R. D., Fairchild, J. F. y Rothermel, B. B. (2004). Effects of an insecticide on amphibians in large-scale experimental ponds. Ecological Applications 14: 685-691.
Brondizio, E., Díaz, S. M., Settele, J., Ngo, H., Guèze, M., Aumeeruddy-Thomas, Y., Bai, X., Geschke, A., Molnár, Z., Niamir, A., Pascual, U., Simcock, A., Jaureguiberry, P., Hien, N., Brancalion, P., Chan, K. M. A., Dubertret, F., Hendry, A., Liu, J., ... Zayas, C. (2019). Assessing a planet in transformation: Rationale and approach of the IPBES Global Assessment on Biodiversity and Ecosystem Services. En IPBES (Ed.), Global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (pp. 5-58). IPBES. https://ri.conicet.gov.ar/handle/11336/151345
Cabrera, M. R. (2015). Reptiles del Centro de la Argentina. Córdoba (Argentina): Editorial de la Universidad Nacional de Córdoba.
Cespedez, J. A., Zaracho, V. H., Álvarez, B. B. y Colombo, M. C. (2008). Diversidad de anfibios: su importancia en los ecosistemas y declinación de poblaciones. En: Manual de Biodiversidad de Chaco, Corrientes y Formosa (pp. 125-135). Córdoba, Argentina: Editorial Universidad Nacional del Nordeste. https://ri.conicet.gov.ar/handle/11336/123058
Collins, E., Castro, S., Felizia, A. y Collins, J. (2022). Estudio de calidad ambiental en la cuenca del arroyo Saladillo, Provincia de Santa Fe. Hacia la obtención de una herramienta de evaluación de calidad. En IFRH 2016. 3er Encuentro de Investigadores en Formación de Recursos Hídricos. Ezeiza, Buenos Aires. https://www.ina.gov.ar/ifrh-2016/trabajos/IFRH_2016_paper_91.pdf
Cushman, S. A. (2006). Effects of habitat loss and fragmentation on amphibians: A review and prospectus. Conservation Biology 128: 231-240. https://doi.org/10.1016/j.biocon.2005.09.031
Cuzziol Boccioni, A. P., Lajmanovich, R.C., Attademo, A. M., Lener, G., Lien-Medrano, C. R., Simoniello, M. F. y Peltzer, P. M. (2024). Toxicity of pesticide cocktails in amphibian larvae: understanding the impact of agricultural activity on aquatic ecosystems in the Salado River basin, Argentina. Drug and Chemical Toxicology 48: 1-19.
Fox, B. y Fox, M. (2000). Factors determining mammal species richness on habitat islands and isolates: habitat diversity, disturbance, species interactions and guild assembly rules. Global Ecology and Biogeography 9: 19-37.
Gallina, S. y López-González, C. (2011). Manual de técnicas para el estudio de la fauna. Volumen I. Querétaro (México): Universidad Autónoma de Querétaro-Instituto de Ecología, A. C.
Gastaudo, J., Anibalini, V. A., Dickie, M. J., Jozami, E., Barbero, S., Pistarelli, D., Fischer, L., Yurun, V., Zarich Icutza, M. B. y Coronel, A. (2022). “La Niña” y la sequía en Zavalla. Revista Agromensajes 64: 32-34.
Ghirardi, R. y López, J. A. (2017). Anfibios de Santa Fe. Santa Fe, Argentina: Ediciones UNL.
Halstead, B. J., Kleeman, P. M., Rose, J. P. y Fellers, G. M. (2022). Sierra Nevada amphibians demonstrate stable occupancy despite precipitation volatility in the early 21st Century. Frontiers in Ecology and Evolution 10: 1040114. https://doi.org/10.3389/fevo.2022.1040114
IBM. (2020). IBM SPSS Statistics for Windows, Version 27.0. Armonk (NY): IBM Corp.
Iriondo, M. H. (2007). The Middle Paraná River: limnology of a subtropical wetland. Berlin: Springer Science & Business Media.
IUCN SSC Amphibian Specialist Group. (2023). Elachistocleis bicolor. The IUCN Red List of Threatened Species 2023: e.T78911125A78910991. https://dx.doi.org/10.2305/IUCN.UK.2023-1.RLTS.T78911125A78910991.en
Jost, L. (2006). Entropy and diversity. Oikos 113(2): 363-375.
Kacoliris, F. (2024). La declinación global de anfibios: Una historia con final incierto. En J. Williams y G. Tettamanti (Eds.), Anfibios de Buenos Aires (pp. 32-35). Buenos Aires: La Biblioteca del Naturalista.
Kacoliris, F. P., Berkunsky, I., Acosta, J. C., Acosta, R., Agostini, M. G., Akmentins, M. S., Arellano, M. L., Azat, C., Bach, N. C., Blanco, G. M., Calvo, R., Charrier, A., Corbalán, V., Correa, C., Cuello, M. E., Deutsch, C., Di Pietro, D. O., Gastón, M. S., Gómez-Alés, R., ... y Williams, J. D. (2022). Current threats faced by amphibian populations in the southern cone of South America. Journal for Nature Conservation 69: 126254. https://doi.org/10.1016/j.jnc.2022.126254
Lajmanovich, R. C., Sánchez-Hernández, J.C., Stringhini, G. y Peltzer, P. M. (2004). Levels of Serum Cholinesterase Activity in the Rococo Toad (Bufo paracnemis) in Agrosystems of Argentina. Bulletin of Environmental Contamination and Toxicology 72(3): 586-591.
Lavilla, E. O. (2001). Amenazas, declinaciones poblacionales y extinciones en anfibios argentinos. Cuadernos de Herpetología 15(1): 59-82.
Lomolino, M. y Perault, D. (2001). Island biogeography and landscape ecology of mammals inhabiting fragmented, temperate rain forests. Global Ecology and Biogeography 1: 113-132.
Lopez, J. A., Scarabotti, P. A. y Ghirardi, R. (2015). Amphibian trophic ecology in increasingly human-altered wetlands. Herpetological Conservation and Biology 10: 819-832.
Magurran, A. y McGill, B. (2011). Biological diversity, frontiers in measurement and assessment. Oxford (UK): Oxford University Press.
Matteucci, S. D. (2012). Ecorregión Pampa. En Ecorregiones y complejos ecosistemicos argentinos. Buenos Aires (Argentina): Orientación Gráfica Editora.
Moreno, C. (2001). Métodos para medir la biodiversidad. Manuales y Tesis SEA, vol. 1. Zaragoza (España): Sociedad Entomológica Aragonesa.
Parris, K. M. (1999). Review: Amphibian survey in forest and woodlands. Contemporary Herpetology 1: 1-20. https://doi.org/10.17161/ch.vi1.11931
Peltzer, P. M., Lajmanovich, R. C., Attademo, A. M. y Cejas, W. (2005). Diversidad y conservación de anuros en ecosistemas agrícolas de Argentina: implicancias en el control biológico de plagas. Serie Miscelánea 14: 399-416.
Peltzer, P. M. (2006). La fragmentación de hábitat y su influencia en la diversidad y distribución de anfibios anuros de áreas ecotonales de los dominios fitogeográficos amazónico y chaqueño. Tesis doctoral. La Plata (Argentina): Universidad Nacional de La Plata.
Perovic, P., Trucco, C., Tálamo, A., Quiroga, V., Ramallo, D., Lacci, A., Baungardner, A. y Mohr, F. (2008). Guía técnica para el monitoreo de la biodiversidad. Salta (Argentina): APN/GEF/BIRF.
Pesci, G. P., Sánchez, J. M., Suelem Muniz, L. y Pelegrin, N. (2018). Reptiles y anfibios de una localidad del Chaco Húmedo en Formosa, Argentina. Cuadernos de Herpetología 32(1): 47-54.
R Core Team. (2025). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.r-project.org/
Raffo, L., Bo, R. y Sanchez, L. (2023). Parque Nacional Ciervo de los Pantanos: Diez años de seguimiento de las comunidades de anfibios anuros. Revista Latinoamericana de Herpetología 6(2): 215-229.
Rodríguez Parra, L. M. (2023). Monitoreo de herpetofauna presente en la Reserva Natural Umbral Cultural Horizontes ubicada en la cadena montañosa de los cerros de Bogotá. Trabajo de grado. Bogotá D.C.: Universidad Distrital Francisco José de Caldas.
Solbrig, O. T. (1999). Bases para una agricultura de altos rendimientos. Nuestro campo Año VII(٦٦).
Suárez, R. P., Zaccagnini, M. E., Babbitt, K. J., Calamari, N. C., Natale, G. S., Cerezo, A., Codugnello, N., Boca, T., Damonete, M. J., Vera-Candioti, J. y Gavier-Pizarro, G. I. (2016). Anuran responses to spatial patterns of agricultural landscapes in Argentina. Landscape Ecology 31: 2485-2505.
Truitt, G. (2019). Monocultivos: la amenaza de los “desiertos verdes” de hoy para la producción alimentaria de mañana. Nature.org. Acceso 22 octubre 2024. https://www.nature.org/es-us/que-hacemos/nuestra-vision/perspectivas/monocultivos-amenaza-desiertos-verdes-produccion-alimentaria/
Ubeda, C. y Grigera, D. (2003). Análisis de la evaluación más reciente del estado de conservación de los anfibios y reptiles de Argentina. Gayana (Concepción) 67(1): 97-113.
Uribe Botero, E. (2015). El cambio climático y sus efectos en la biodiversidad en América Latina. Santiago de Chile: Comisión Económica para América Latina y el Caribe (CEPAL).
Vaira, M., Akmentins, M. S., Attademo, M., Baldo, D., Barrasso, D., Barrionuevo, S., Basso, N., Blotto, B., Cairo, S., Cajade, R., Céspedez, J., Corbalán, V., Chilote, P., Duré, M., Falcione, C., Ferraro, D., Gutierrez, R., Ingaramo, M., Junges, C., ... Zaracho, V. (2012). Categorización del estado de conservación de los anfibios de la República Argentina. Cuadernos de Herpetología 26(Supl. 1): 131-159.
Valencia-Aguilar, A., Cortés-Gómez, A. M. y Ruiz-Agudelo, C. A. (2012). Servicios ecosistémicos brindados por los anfibios y reptiles del Neotrópico: una visión general. Reflexiones sobre el capital natural de Colombia 2: 1-26.
Veloza, F. (2020). Diversidad funcional y taxonómica de la herpetofauna en la Subcuenca Bajo Río Bogotá, en el departamento de Cundinamarca. Tesis de maestría. Bogotá: Pontificia Universidad Javeriana.
Vera, D. G., Kacoliris, F. P. y Williams, J. D. (2021). Ecología espacial y prioridades de conservación en reptiles nativos que habitan relictos de pastizal del sistema serrano de Tandilia. Investigación Joven 7(2): 416-417.
Volpedo, A. V. y Reyes, L. F. (2008). Efecto de los cambios globales sobre la biodiversidad. La Habana, Cuba: RED CYTED Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo.
Walls, S. C., Barichivich, W. J. y Brown, M. E. (2013). Drought, deluge and declines: The impact of precipitation extremes on amphibians in a changing climate. Biology 2(1): 399-418. https://doi.org/10.3390/biology2010399
Wells, K. D. (2007). The ecology and behavior of amphibians. Chicago: University of Chicago Press.
Williams, J.D., Tettamanti, G., Vera, D.G., Baguette Pereiro, B., Prosdocimi, L., Grilli, P. G. y Kacoliris, F. G. (2022). Reptiles de Buenos Aires. Buenos Aires: La Biblioteca del Naturalista.
Williams, J.D. y Tettamanti, G. (2024). Anfibios de Buenos Aires. Buenos Aires: La Biblioteca del Naturalista.