Articulo original
Original article
36 (1) | Ene - Jun, 2026
Facultad de Ciencias Exactas y Naturales y Agrimensura (UNNE)
Open Access: https://revistas.unne.edu.ar/index.php/fce
E-mail: revistafacena@exa.unne.edu.ar
Secuencia de condrificación y osificación postembrionaria en Physalaemus biligonigerus (Anura: Leptodactylidae)
Sequence of postembryonic chondrification and ossification in Physalaemus biligonigerus (Anura: Leptodactylidae)
Stechina, Ornela Sofía ¹ y Zaracho, Victor Hugo * ²
1. Laboratorio de Biología Molecular (Instituto de Medicina Regional, Universidad Nacional del Nordeste) Resistencia, Argentina.
2. Laboratorio de Herpetología de la Facultad de Ciencias Exactas y Naturales y Agrimensura (Universidad Nacional del Nordeste). Corrientes, Argentina.
* Autor de correspodencia: victorzaracho@exa.unne.edu.ar
Recibido/Received: 11 de febrero, 2026 | Aceptado/Acepted: 24 de junio, 2026 | Publicado/Published: 30 de junio, 2026
Como citar este artículo: Stechina, O. S. & Zaracho, V. H. (2026). Secuencia de condrificación y osificación postembrionaria en Physalaemus biligonigerus (Anura: Leptodactylidae). Revista FACENA 36(1), 110-139. Doi: https://doi.org/10.30972/fac.3619117
Resumen: Los caracteres larvales, en especial los osteológicos, son una fuente valiosa para estudios taxonómicos, filogenéticos, y comparativos en anuros. Aunque la ontogenia postembrionaria es aún poco explorada, los enfoques evo-devo muestran que las trayectorias de desarrollo revelan mecanismos de diversificación biológica, con la heterocronía como proceso central. En este sentido, las secuencias de condrificación y osificación de anuros puede contribuir a identificar desplazamientos temporales y patrones evolutivos. Aquí describimos la secuencia postembrionaria de condrificación y osificación de Physalaemus biligonigerus, en comparación con otros Leptodactylidae. Se analizaron larvas entre los estadios 26-46 de Gosner, diafanizadas y teñidas diferencialmente para huesos y cartílagos. En P. biligonigerus, los primeros huesos en osificarse son el frontoparietal, el paraesfenoides y los exoccipitales. En el esqueleto postcraneal, la osificación se inicia con los huesos largos, siendo las falanges los últimos en osificarse. Hacia el estadio 46 la supraescápula, el procoracoides, el epiesternón, el xifiesternón, los carpales, el extremo distal del ilion, gran parte del isquion, el pubis, y los tarsales permanecen cartilaginosos. Si bien el patrón de condrificación y osificación de P. biligonigerus es similar al de otros Leptodactylidae, presenta particularidades en el tiempo de aparición y en el grado de desarrollo de diversos elementos craneales y postcraneales. Las diferencias con especies cercanas evidencian la diversidad de trayectorias ontogenéticas dentro del clado, y resaltan la necesidad de comparar secuencias de diferenciación, un enfoque que permite evaluar el orden relativo de aparición de estructuras sin depender de estadios susceptibles a variación heterocrónica. Desde una perspectiva evo-devo, los desplazamientos temporales observados reflejan procesos heterocrónicos que modulan la maduración de distintos módulos esqueléticos y contribuyen a la diversidad morfológica dentro del clado. En conjunto, los patrones documentados para P. biligonigerus podrían constituir una base comparativa sólida para futuros estudios sobre heterocronía, evolución morfológica y relaciones filogenéticas en anuros.
Palabras claves: Ontogenia; Larvas; Cráneo; Postcráneo.
Abstract: Larval characters, particularly osteological ones, are a valuable source of evidence for taxonomic, phylogenetic, and comparative studies in anurnas. Although post-embryonic ontogeny remains poorly explored, evodevo approaches show that developmental trajectories reveal mechanisms of biological diversification, with heterochrony playing a central role. In this context, anuran chondrification and ossification sequences can help identify temporal shifts and evolutionary patterns. Here we describe the postembryonic chondrification and ossification sequence of Physalaemus biligonigerus, in comparison with other Leptodactylidae. Larvae between Gosner stages 26–46 were cleared and differentially stained for bones and cartilages. In P. biligonigerus, the first bones to ossify are the frontoparietal, parasphenoid, and exoccipitals. In the postcranial skeleton, ossification begins with the long bones, while the phalanges are the last to ossify. By stage 46, the suprascapula, procoracoid, epi and xiphisternum, carpals, distal end of the ilium, most of the ischium, the pubis, and the tarsals remain cartilaginous. Although the chondrification and ossification pattern of P. biligonigerus is broadly similar to that of other Leptodactylidae, it shows particularities in the timing and degree of development of several cranial and postcranial elements. Differences with closely related species highlight the diversity of ontogenetic trajectories within the clade and underscore the value of comparing differentiation sequences, an approach that allows evaluating the relative order of appearance of structures without relying on stages susceptible to heterochronic variation. From an evodevo perspective, the temporal shifts observed reflect heterochronic processes that modulate the maturation of different skeletal modules and contribute to morphological diversity within the clade. Taken together, the patterns documented for P. biligonigerus may provide a useful comparative basis for future studies on heterochrony, morphological evolution, and phylogenetic relationships in anurans.
Keywords: Ontogeny; Tadpoles; Skull; Postcranium.
Introducción
El desarrollo embrionario y larval de anuros ha sido caracterizado tradicionalmente a partir de los cambios observables en la morfología externa, y muchos de ellos se encuentran sistematizados en diversas tablas de desarrollo. Entre ellas, la propuesta clásica de Gosner (1960), compuesta por 46 estadios, continúa siendo una referencia ampliamente utilizada y aplicada a muchas familias de anuros, más allá que fue diseñada originalmente para Bufonidae, Hylidae, Pelobatidae y Ranidae de América del Norte. Paralelamente, los caracteres osteológicos, y en particular los derivados del esqueleto larval han demostrado ser una fuente valiosa de información para estudios taxonómicos, filogenéticos, y comparativos (Larson y de Sá 1998; Haas, 2003). La mayoría de los estudios disponibles se han centrado en descripciones del condrocráneo y del esqueleto visceral en estadios específicos del desarrollo. En contraste, estudios que aborden los patrones del desarrollo postembrionario en un marco comparativo amplio, o en contexto filogenético son todavía limitados (Maglia et al., 2001).
En las últimas décadas, los enfoques de la biología del desarrollo evolutivo (evo-devo) han resaltado la importancia de los procesos morfogenéticos como generadores de variación evolutiva, enfatizando que las trayectorias ontogenéticas pueden revelar mecanismos que subyacen a la diversificación morfológica (Hall, 2012; Müller, 2007). En este contexto, la heterocronía, definida como cambios en el tiempo o en la tasa de desarrollo de estructuras durante la ontogenia, constituye un proceso clave para interpretar la evolución de los patrones esqueléticos en vertebrados (Smith, 2003). En anuros, las secuencias de condrificación y osificación han permitido identificar desplazamientos temporales entre taxones y reconocer tendencias evolutivas y establecer patrones generales del desarrollo esquelético, constituyendo además una herramienta relevante para la interpretación de la diversidad morfológica (Hanken y Hall, 1988; Haas, 1999). Estudios recientes han reforzado este enfoque al destacar la modularidad e integración del esqueleto de los anuros como componentes esenciales para comprender la evolución del desarrollo (Klingenberg, 2010; Kuratani, 2009; Fabrezi et al., 2017a).
Los antecedentes disponibles sobre patrones de condrificación y osificación en anuros abarcan un amplio espectro de linajes, entre ellos Hylidae (Stokely y List, 1954; de Sá, 1988; Freemann y de Sá, 1995; Arenas-Rodriguez et al., 2018; Fabrezi y Goldberg, 2009; Rynophrynidae (Trueb, 1985); Pelodytidae (Talavera, 1985); Scaphiopodidae (Wiens, 1989; Banburi y Maglia, 2006; Hall y Larsen Jr., 1998); Microhylidae (de Sá y Trueb, 1991); Phyllomedusidae (de Sá y Lavilla, 1996; Sheil y Alamillo 2005); Bombinatoridae (Maglia y Pugener 1998); Pyxicephalidae (Haas, 1999; Sheil, 1999); Rhacophoridae (Kerney et al., 2007; Senevirathne et al., 2017); Alsodidae (Vera Candioti et al., 2005); Pipidae (Trueb y Hanken, 1992; Trueb et al., 2000); Ceratophryidae (Wild, 1997; Wild, 1999); Bufonidae (Yıldırım y Kaya, 2016; Kovalenko y Kruzhkova, 2013; Gómez et al., 2017); Alytidae (Púgener y Maglia, 1997) y Dendrobatidae (de Sa y Hill, 1998). En Leptodactylidae, los aportes son limitados y se restringen a algunas especies como Leptodactylus macrosternum (Perotti, 2001), L. bufonius (Vera y Ponssa, 2013) y L. latinasus (Gómez et al., 2017).
En un marco comparativo más amplio, diversos estudios han abordado aspectos generales del desarrollo esquelético, como la formación de exoccipitales, paraesfenoides y frontoparietales durante la metamorfosis (Hanken y Hall, 1988), los patrones de diversidad craneal en Lissamphibia (Trueb, 1992), el desarrollo comparado en un contexto filogenético (Maglia et al. 2001), los patrones del desarrollo carpal (Fabrezi y Barg, 2001), la variación del autopodio en anuros (Goldberg y Fabrezi, 2008), y la ontogenia de la cintura pélvica y las extremidades posteriores en anuros (Manzano et al., ٢٠١٣)
Physalaemus Fitzinger, 1826 incluye unas 50 especies de ranas sudamericanas que, junto a los géneros Edalorhina, Engystomops, Pleurodema y Pseudopaludicola conforman el clado Leiuperinae (Frost, 2025). De acuerdo a estudios filogenéticos recientes, se reconocen dos clados principales, entre los que se encuentra el clado de Physalaemus cuvieri, al cual pertenece la especie en estudio, P. biligonigerus (Cope, 1861) (Lourenco et al., 2015), y cuya distribución comprende la región norte y centro de Argentina, Bolivia, Paraguay, Uruguay y el sur de Brasil (Frost, 2025).
Si bien existen descripciones del condrocráneo para algunas especies de Physalaemus, entre ellas de P. biligonigerus (Fabrezi y Vera, 1997), los patrones del desarrollo esquelético en este género permanecen prácticamente desconocidos. En este contexto, la comparación de secuencias de desarrollo osteológico entre Physalaemus y otros géneros de Leptodactylidae permitirá incorporar evidencia novedosa para el género, identificar procesos heterocrónicos y comprender patrones y trayectorias de desarrollo dentro del grupo.
El presente trabajo describe la secuencia de condrificación y osificación larval de Physalaemus biligonigerus y caracteriza los principales elementos condrales y óseos del esqueleto craneal y postcraneal durante el desarrollo postembrionario. Asimismo, se compara con la de otros leptodactílidos con el fin de identificar similitudes y diferencias en los patrones de desarrollo, reconocer posibles cambios heterocrónicos y contribuir a la comprensión de las trayectorias ontogenéticas dentro de la familia.
MATERIALES Y MÉTODOS
Se utilizaron series larvales de Physalaemus biligonigerus depositadas en la Colección Herpetológica de la Universidad Nacional del Nordeste (UNNEC 14473), procedentes de puestas coleccionadas en charcas temporarias de la localidad de Comandante Fontana (Patiño, Formosa, Argentina). Los estadios larvales fueron identificados empleando la Tabla de Gosner (1960) bajo una lupa binocular Leica Zoom 2000. Los estadios considerados incluyen desde el 26 al 46 y, en general, se examinaron de 3 a 5 individuos por estadio, de acuerdo con la disponibilidad del material (se priorizaron ejemplares que presentaron un buen estado de conservación y una tinción diferencial adecuada para la identificación de los elementos condrales y óseos). Para la descripción, los estadios fueron agrupados de a pares y, a su vez, en rangos más amplios, dado que entre algunos estadios consecutivos no se observaron cambios significativos. No obstante, cuando fue necesario resaltar eventos particulares del desarrollo, se conservó la identidad de los estadios individuales. Para la diferenciación de los elementos condrales y óseos se empleó la técnica de diafanización y tinción diferencial de hueso y cartílago, mediante la cual los tejidos cartilaginosos se tiñen de azul y los tejidos óseos de rojo (Wassersug, 1976).
La terminología empleada sigue a Lavilla y Fabrezi (1987), Fabrezi y Lavilla (1993) y Larson y De Sa (1998). Para la representación gráfica se confeccionaron dibujos con cámara clara utilizando un microscopio estereoscópico Olympus SZX9 y se obtuvieron fotografías con una lupa Leica EZ4 equipada con cámara integrada.
resultados
Estadios 26-31. Durante este periodo están presentes únicamente componentes del esqueleto axial (condrocráneo, esqueleto hiobranquial y arcos neurales), todos cartilaginosos. En estadios 26-27 (Figuras 1A y 2A), el condrocráneo es relativamente más largo que ancho; los cuernos trabeculares son planos y divergentes, equivalentes al 23% de la longitud total del condrocráneo. La fenestra basicraneal tiene forma ovalada y representa alrededor del 36% de la longitud total del condrocráneo; a finales de esta etapa comienza a cerrarse en su región anterior, y en la región posterior, aparecen esbozos de los forámenes carotídeos. Las cápsulas óticas tienen aspecto cuadrangular y representan aproximadamente el 29% de la longitud total. A partir del estadio 28-29 son distintivas las crestas paróticas, y algunas larvas, en estadio 30-31, ya presentan un techo sinótico débilmente condrificado. Los arcos occipitales están en continuidad con el planun basale y definen los cóndilos occipitales. Los forámenes yugular y perilinfático inferior son distintivos. En la cara ventral de las cápsulas óticas se distinguen los forámenes perilinfáticos. El proceso ascendente del palatocuadrado es de unión baja con el neurocráneo. Una comisura cuadrado-orbital está presente, por lo cual el canal muscular está cerrado. El cartílago de Meckel tiene forma de “S”, y presenta un proceso retroarticular bien diferenciado. El suprarrostral está débilmente condrificado en los estadios 26-27, y desde el estadio 28-29 adquiere su configuración tetrapartita. En el estadio 30-31 (Figuras 1B y 2B) se diferencian los procesos laterales de los cuernos trabeculares, el esbozo del tectum nasi y la cresta parótica presenta un proceso anterolateral distintivo. Esqueleto hiobranquial: pars reuniens y ceratohiales bien condrificados desde los estadios 26-27 (Figura 3A); ceratobranquiales con diferentes grados de desarrollo. En los estadios 28-29 la pars reuniens se fusiona con la cópula II y el ceratohial. En los estadios 30-31 (Figura 3B) los procesos branquiales de los ceratobranquiales II y III están diferenciados, pero no entran en contacto (abiertos), y los ceratobranquiales I y II están fusionados a la placa hipobranquial. El esqueleto axial aparece en los estadios 28-29, representados por 5-6 esbozos pares de arcos neurales (Figura 4A), que aumentan a 9 pares en los estadios 30-31 (Figura 4B). Durante esta etapa no se observan elementos del esqueleto apendicular.
Estadio 32-37. Durante estos estadios continúa la condrificación del condrocráneo y aparecen los primeros elementos del esqueleto apendicular. En el condrocráneo, a partir del estadio 32-33 comienza a cerrarse la fenestra basicraneal desde su región anterior, mientras en la región posterolateral quedan definidos los forámenes carotídeos, delimitados totalmente por cartílago. Se observa además el techo sinótico, completamente condrificado. En los estadios 34-35, se observan en la región posteroventral de la pared lateral, los forámenes oculomotores, delimitados por una delgada franja de cartílago (cartílagos orbitales). Hacia el estadio 36-37 la fenestra basicraneal queda reducida a un tercio de su tamaño original. Esqueleto hiobranquial: sin cambios significativos (Figura 3C). Esqueleto axial: al inicio se observan los 9 arcos previamente mencionados (Fig. 4C), pero hacia finales de esta etapa se diferencian 10 arcos neurales; el último queda reducido a una pieza triangular que contribuirá a la formación del urostilo (Figura 4D). Las postzigapófisis de las vértebras II-VIII y las apófisis transversas de las vértebras II-III están bien desarrolladas.
Con respecto al esqueleto apendicular (Figuras 5 y 6), en el estadio 32-33 se observan los primeros esbozos de la extremidad posterior (fémur, tibia y fíbula) (Figura 6A). Hacia el estadio 34/35 estos componentes están más alargados, aparecen esbozos de los tarsales proximales, metatarsal III, y un esbozo de la cintura pélvica (Figura 6B); en la extremidad anterior se esbozan el húmero y elementos del zeugopodio. En el estadio 36-37, la cintura pectoral está formada por tres esbozos cartilaginosos: procoracoides y coracoides, ventrales, paralelos y con forma de espinas, y la escápula, dorsal, y con una ligera constricción en la parte media; el húmero es robusto y el radio y la ulna se presentan como cartílagos independientes; en el carpo, se diferencian el radial, ulnar y carpal distal III-V, y el metacarpal proximal del IV dedo también está presente (Figura 5A). La cintura pélvica tiene forma de gancho con el ilion y el isquion bien diferenciados; no se observan estructuras cartilaginosas que puedan asociarse con el pubis; en la extremidad posterior están presentes el fémur, tibia, fíbula, el tibial y fibular como cartílagos independientes, junto a los metatarsales II-V y la falange proximal del IV (Figura 6C).
Estadio 38-41.
Estos estadios están marcados por el inicio de la osificación y la diferenciación de nuevas estructuras craneales. En el condrocráneo la fenestra basicraneal está completamente cerrada, aunque débilmente condrificada en su parte central. Los forámenes craneopalatinos están ahora bien delimitados. A nivel de las cápsulas óticas se observan algunas calcificaciones internas. Aparece el esbozo de septum nasi y los cartílagos orbitales siguen limitados a un estrecho puente cartilaginoso sobre el foramen oculomotor. Hacia el estadio 40-41 (Figura 1D) se observa el septum nasi como una lámina sagital, estrecha y alta, mientras que el tectum nasi se esboza como dos proyecciones cartilaginosas orientadas en V; se esbozan además las taeniae tecti transversalis y medialis. El frontoparietal inicia su osificación sobre la fontanela frontoparietal, constituyendo el primer hueso en formarse. Esqueleto hiobranquial: sin cambios significativos (Figura 3D).
Esqueleto axial: en este periodo (Figura 4 E-G) los cuerpos vertebrales presentan los primeros esbozos de condrificación, en tanto los arcos neurales progresan en su cierre dorsal: los arcos I-II quedan próximos entre sí, los III-V cerrados y los VI-IX en contacto; las prezigapófisis y postzigapófisis de todas las vértebras están en contacto con sus vértebras contiguas (Figura 4F). El arco neural X es rudimentario y aparece el esbozo de la hipocorda (Figura 4H).
Esqueleto apendicular (Figura 5 y 6): a partir de estadio 38-39 comienzan a aparecer los primeros centros de osificación en la región media de la diáfisis de todos los huesos largos, constituyendo el hito más importante de este intervalo. En la cintura pectoral se distinguen la pars acromialis y la pars glenoidea en la escápula, y el procoracoides y coracoides continúan con forma de espina; se observa además la supraescápula y la clavícula (Figura 5B). El radio y la ulna se fusionan en su extremo distal. En el carpo (Figura 5C) se identifican cinco piezas cartilaginosas: ulnar, radial, carpal distal III-V, carpal distal II, y el elemento Y. El prepollex está representado por dos piezas cartilaginosas. Se diferencian los metacarpales de los cuatro dedos y la mayoría de las falanges. Se registran los primeros centros de osificación en la escápula, en el húmero, en el radio y en la ulna. En la cintura pélvica se diferencia el pubis, cartilaginoso (Figura 6D). La tibia y la fíbula están en contacto, excepto en su parte media. El tibial y fibular se fusionan en sus extremos. A finales de esta fase (Figura 6E), en el tarso se observa el tarsal distal II-III, el elemento Y, y el prehallux (representado por una única pieza). Todos los metatarsales están presentes, igual que las falanges, excepto la falange distal del dedo I y del dedo V. Se registran los primeros centros de osificación en el ilion, en el fémur, en la tibia, en la fíbula, y en los metatarsales I-IV.
Estadio 42-46.
Estos estadios comprenden el clímax metamórfico y están caracterizados por una osificación progresiva y generalizada del esqueleto craneal y postcraneal, junto con la remodelación de varias estructuras larvales.
Cráneo: al inicio de esta fase el palatocuadrado muestra algunos cambios en su conformación: el proceso ascendente degenera y pierde la conexión con el neurocráneo y el proceso muscular se desplaza hacia la región media del ojo; el paraesfenoides está osificado, presenta un proceso cultriforme largo que alcanza el extremo anterior de las órbitas, y los procesos alares se extienden hacia las cápsulas óticas; los exoccipitales presentan las primeras evidencias de osificación, mientras que el basioccipital y el supraoccipital permanecen cartilaginosos (Figura 1E, 2E). Hacia el estadio 44 (Figura 1F, 2F) los frontoparietales están osificados y separados en la línea media; los nasales, osificados, son subtriangulares; los premaxilares, sin dientes, están osificados; el escamoso se observa como una varilla muy delgada sobre parte del palatocuadrado; el proceso proótico está osificado en su región anterior. En el estadio 46 (Figura 1G, 2G), se observan dientes en el premaxilar, y el maxilar está osificado en su extremo anterior, y también presenta dientes; el septomaxilar está osificado, el escamoso es claramente distintivo con forma de espina, y la osificación en los exooccipitales se extiende dorsal y ventralmente; el dentario presenta esbozo de osificación en su extremo posterior.
Esqueleto hiobranquial: hacia el estadio 42/43 los ceratobranquiales II-IV se reducen y desaparecen el proceso anterolateral del ceratohial, los radios y las espículas (Figura 3E). En el estadio 44 desaparecen los ceratobranquiales (Figura 3F). Se observan los cartílagos que darán origen a la laringe. En el estadio 46 el esqueleto hiobranquial adquiere la forma adulta, manteniendo todos sus elementos cartilaginosos (Figura 3G).
Esqueleto axial: los centros vertebrales se osifican débilmente y los arcos neurales se osifican, excepto en la zona de contacto con los cuerpos vertebrales y de la futura espina neural. Las primeras tres apófisis transversas están osificadas en su mayor parte, excepto en sus extremos (Figura 4E-F); la vértebra sacra mantiene sus apófisis transversas cartilaginosas. Los arcos neurales X se fusionan entre sí y contactan con el cuerpo vertebral sacro, con su vértice prolongado posteriormente (Figura 4K). Hacia el estadio 44 (Figura 4L) la columna vertebral está casi totalmente osificada, diferenciándose claramente una vértebra cervical, siete presacras y una sacra; las apófisis transversas están osificadas con sus extremos cartilaginosos y la apófisis transversa sacra permanece cartilaginosa. La hipocorda está osificada en su mitad anterior y en el estadio 46 (Figura 4 M-P) queda asociada a los dos centros de osificación derivados del arco neural X.
Esqueleto apendicular: en el estadio 42-43 la escápula y el coracoides están osificados en su parte media; la supraescápula, el procoracoides y el epicoracoides permanecen cartilaginosos. La clavícula y el cleitro están débilmente osificados (Figura 5D). En la extremidad anterior (Figura 5 E) se observan osificaciones, en diferentes grados, en la parte media de la diáfisis del húmero, radio, ulna, y de algunos metacarpianos. El húmero presenta una cresta humeral distintiva. Los carpales y la mayoría de las falanges permanecen cartilaginosas (Figura 5 F). El prepollex está formado por tres piezas (Figura 5G). En el esqueleto apendicular posterior (Figura 6F), el ilion, con una cresta dorsal bien diferenciada, presenta diferentes grados de osificación, en algunos ejemplares hasta ⅔ de su longitud, manteniéndose cartilaginoso su extremo anterior; el isquion y el pubis son cartilaginosos. El fémur, tibia, fíbula, tibial y fibular presentan osificación avanzada excepto en sus epífisis; los metatarsales III al V están osificados a nivel del tercio medio de sus diáfisis, mientras las falanges se conservan cartilaginosas, con excepción un centro de osificación en el tercio medio de la diáfisis de la falange del dedo IV. El tarso está formado por tres piezas cartilaginosas: el tarsal distal III-II, el tarsal distal I y el elemento Y, estos últimos muy próximos entre sí. El prehallux está compuesto por dos elementos cartilaginosos.
Hacia el estadio 44 el húmero y el radio-ulna están prácticamente osificados en su totalidad; avanza la osificación del cleitro y de la clavícula; los coracoides están parcialmente osificados, mientras que el procoracoides y el epicoracoides permanecen cartilaginosos (Figura 5H). Aparece un centro de osificación en el isquion y los huesos del miembro posterior están completamente osificados excepto sus epífisis, que se mantienen cartilaginosas (Figura 6 G-H). En el estadio 46 (Figura 5 I-J y 6 M-P), se conservan cartilaginosos la supraescápula, el procoracoides, el epiesternón, el xifiesternón, los carpales, el extremo anterior del ilion, gran parte del isquion, el pubis, y los tarsales.
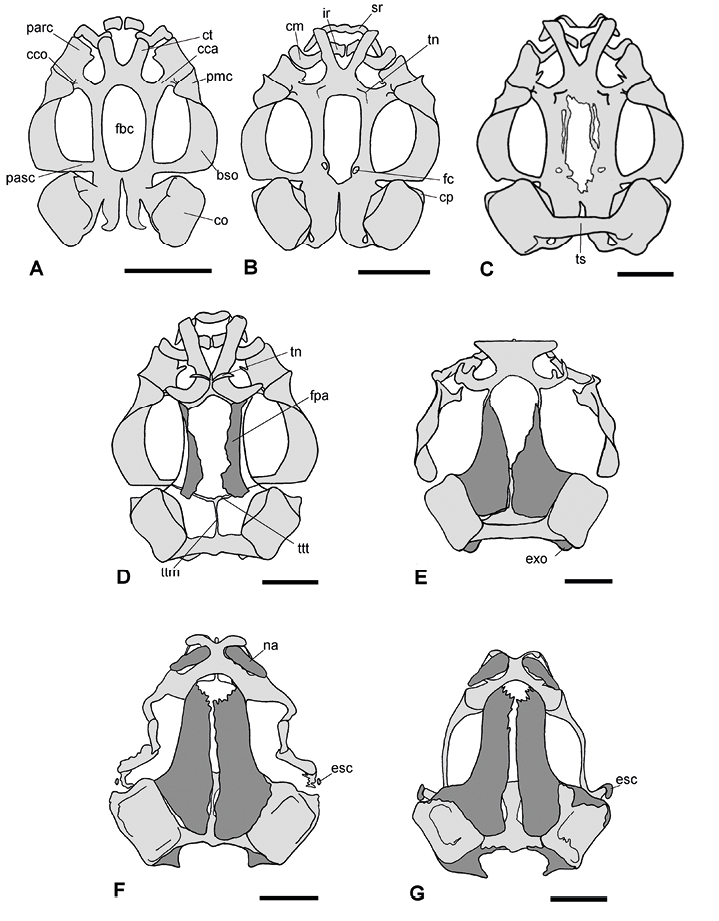
Figura 1. Secuencia ontogenética del desarrollo craneal en vista dorsal de Physalaemus biligonigerus. Las regiones correspondientes a tejido cartilaginoso se representan en gris claro, y las correspondientes a tejido óseo, en gris oscuro. A, estadio 26/27 (condrocráneo completamente cartilaginoso); B, estadio 30/31 (procesos laterales de los cuernos trabeculares presentes y esbozos del tectum nasi); C, estadio 34/35 (comienza el cierre de la fenestra basicraneal y es evidente el techo sinótico); D, estadio 40/41 (frontoparietales creciendo sobre la fontanela frontoparietal, esbozos de las tenias tectis); E, estadio 42/43 (proceso ascendente del palatocuadrado ausente, primeras evidencias de osificación de los exoccipitales); F, estadio 44 (nasales osificados, subtriangulares y separados entre sí por el septo nasal, esbozo del escamoso); G, estadio 46 (supraoccipital comienza a osificarse, escamoso en formación). Escala: 1 mm. Referencias: bso, barra subocular del palatocuadrado; cca, comisura cuadrado craneal anterior; cco, comisura cuadrado orbital; cm, cartílago de Meckel; co, cápsula ótica, cp, cresta parótica; ct, cuernos trabeculares; esc, escamoso; exo, exoocipital; fbc, fenestra basicraneal; fc, foramen carotídeo; fpa, frontoparietal; ir, infrarrostral; tn, tectum nasi; na, nasal; parc, proceso articular del cuadrado; pasc, proceso ascendente el cuadrado; pmc, proceso muscular del cuadrado; sr, suprarrostral; ts, techo sinóptico; tn, tectum nasi; ttt, taenia tecti marginalis; ttm; taenia tecti medialis.
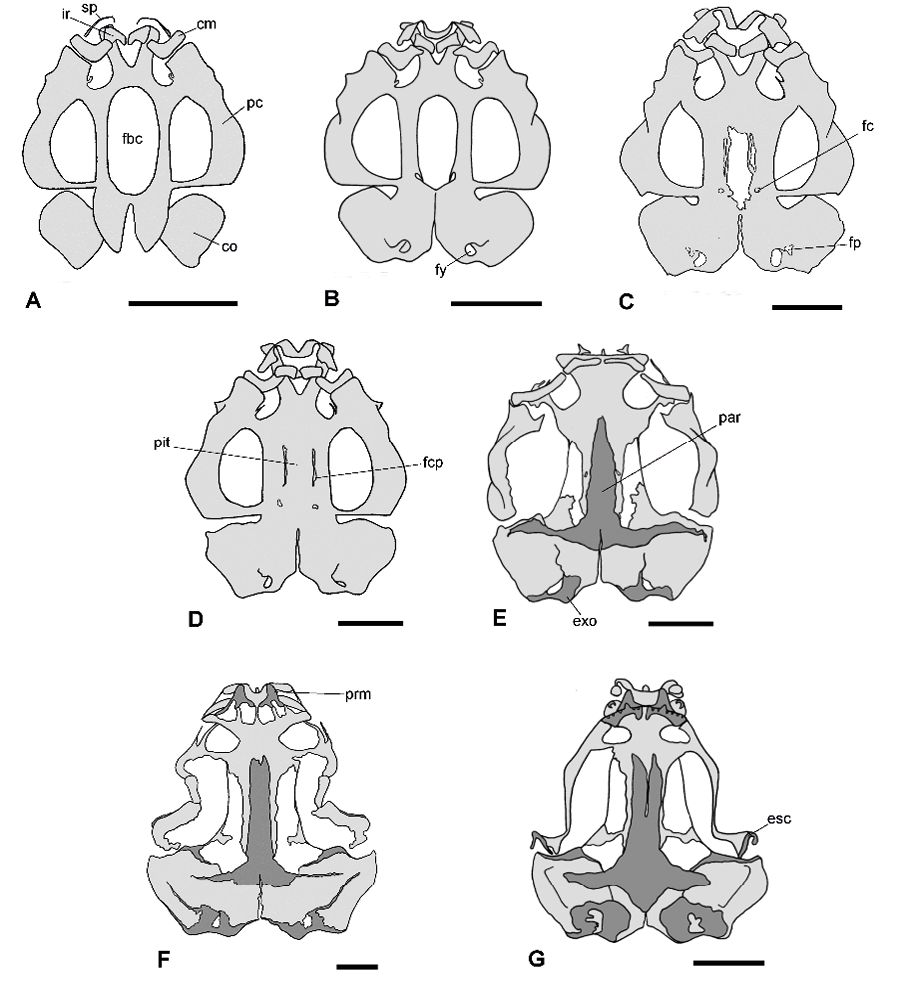
Figura 2. Secuencia ontogenética del desarrollo craneal en vista ventral de Physalaemus biligonigerus. Las regiones correspondientes a tejido cartilaginoso se representan en gris claro, y las correspondientes a tejido óseo, en gris oscuro. A, estadio 26/27 (presencia del cartílago de Meckel e infrarrostrales); B, estadio 30/31 (suprarrostral condrificado y arco occipital bien desarrollado); C, estadio 34/35 (inicio del cierre de la fenestra basicraneal); D, estadio 40/41 (cierre de la fenestra basicraneal); E, estadio 42/43 (paraesfenoides con proceso cultriforme y procesos alares); F, estadio 44 (premaxilares edentados, región anterior del proótico osificado); G, estadio 46 (premaxilares dentados, maxilares edentados, dentario y escamosos osificados). Escala: 1 mm. Referencias: cm, cartílago de Meckel; co, cápsula ótica; esc, escamoso; exo, exoocipital; fbc, fenestra basicraneal; fc, foramen carotídeo; fcp, foramen craneopalatino; fp, foramen perilinfático; fy, foramen yugular; ir, infrarrostral; par, paraesfenoides; pc, palotocuadrado; pit, planum intertrabecular; prm, premaxilar; sr, suprarrostral.
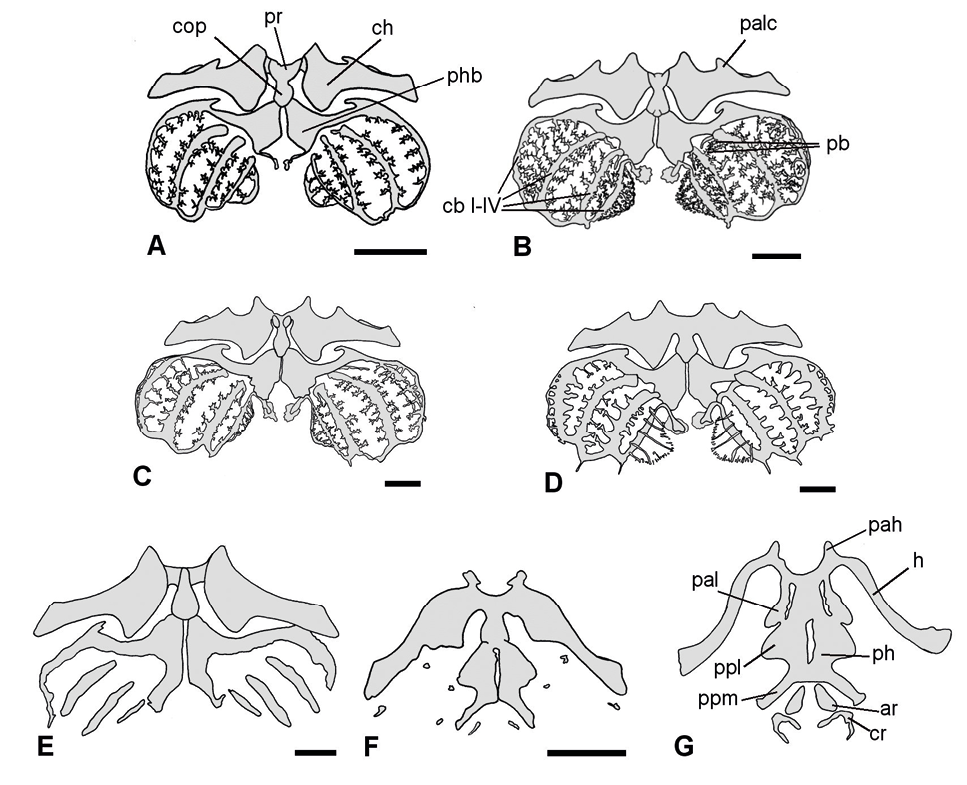
Figura 3. Transformaciones del aparato hioideo durante el desarrollo larval de Physalaemus biligonigerus (vista ventral). A, estadio 26/27; B, estadio 30/31 (procesos branquiales de los ceratobranquiales II y III diferenciados, sin contacto); C, estadio 34/35. D, estadio 40/41 (note la reducción de radios y espículas de los ceratobranquiales); E, estadio 42/43 (reducción de los ceratobranquiales); F, estadio 44 (pérdida de ceratobranquiales). G, estadio 46 (forma adulta, sin evidencias de osificaciones). Escala: 1 mm. Referencias: ar, aritenoides; cb I–IV, ceratobranquiales, ch, ceratohial; cop, cópula II; cr, cricoides; h, hial; pac, proceso anterolateral del ceratohial; pal, proceso anterolateral; pah, proceso anterior; pb, procesos branquiales; ph, placa hioidea; phb, placa hipobranquial; ppl, proceso posterolateral; ppm, proceso posteromedial; pr, pars reuniens.
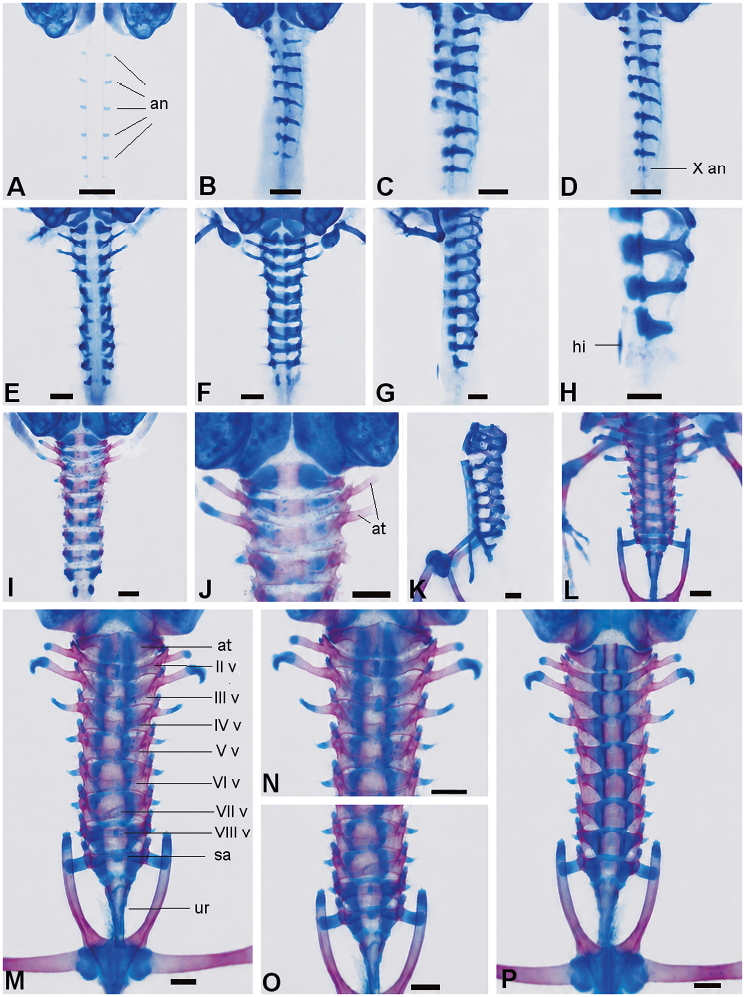
Figura 4. Secuencia de desarrollo del esqueleto axial de Physalaemus biligonigerus (los elementos condrales están representados en azul y los elementos óseos, en rojo. A, Estadio 28/29 (vista ventrolateral, las flechas indican esbozos de arcos neurales); B, estadio 30/31 (vista ventrolateral, note 9 pares de arcos neurales); C, estadio 32/33; D, estadio 37 (la flecha indica el X arco neural); E, estadio 38/39; F-H, estadio 38/39 (F, vista dorsal; G, vista lateral; H, detalle de la región posterior del esqueleto axial mostrado en G, la flecha indica la hipocorda); I-K, estadio 42/43, primeras osificaciones vertebrales (J, detalle de la región anterior mostrando osificaciones de los arcos neurales y apófisis transversas; K, vista lateral, la flecha indica el urostilo en formación); L, estadio 44 (vista dorsal, las vértebras están prácticamente osificadas en su totalidad, note las apófisis transversas de la vértebra sacra cartilaginosa); M-P, estadio 46 (M, vista dorsal completa de la columna vertebral; N, detalle de la región anterior; O, detalle de la región posterior; P, vista ventral completa de la columna vertebral). Escala: 0,5 mm. Referencias: an, arco neural; at, atlas; at, apófisis transversa; hi, hipocorda; sa, sacro; ur, urostilo; v, vértebra.
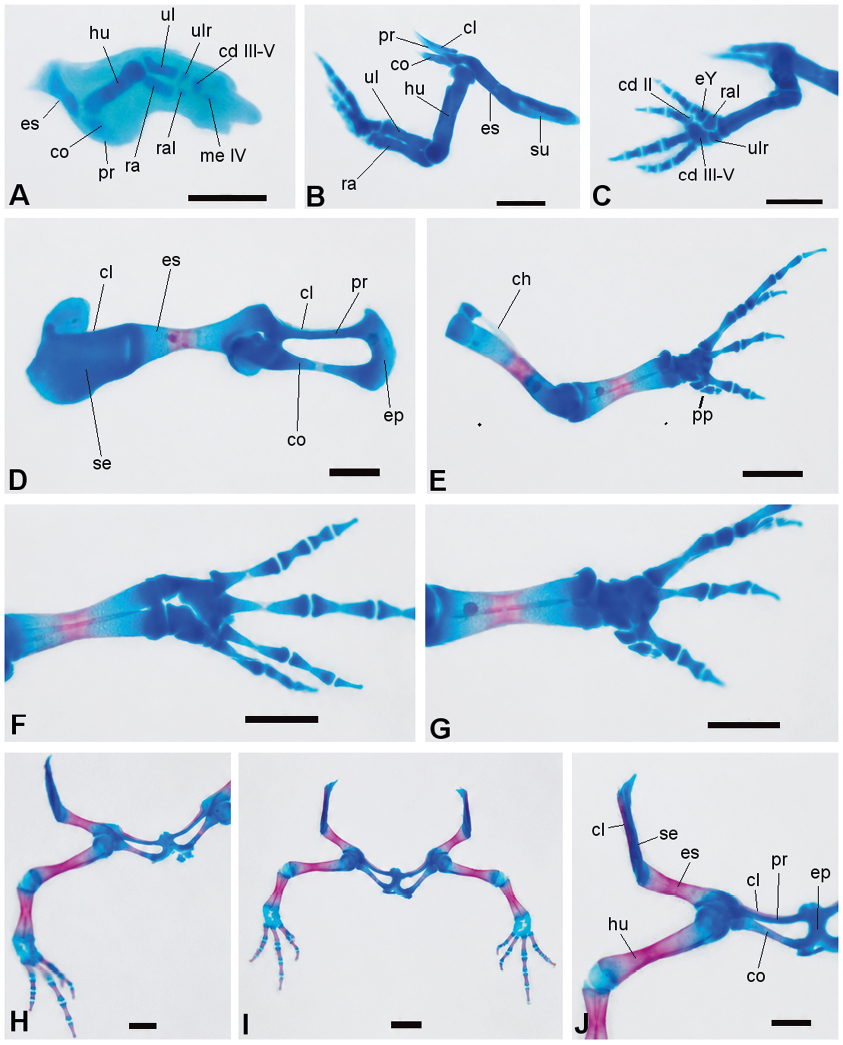
Figura 5. Secuencia de desarrollo de los miembros anteriores y cintura pectoral de Physalaemus biligonigerus (los elementos condrales están representados en azul y los elementos óseos, en rojo. A, estadio 36/37 (en la cintura pectoral, esbozos de escápula, procoracoides y coracoides, en la extremidad anterior, húmero, radio, ulna, y algunos elementos carpales y metacarpales); B-C, estadio 38/39 (B, supraescápula cartilaginosa y clavícula ósea distintivas; C, note los elementos del autopodio); D-G, estadio 42/43, note el avance de las osificaciones (D, cintura pectoral; E, extremidad anterior; F-G, detalle del autopodio); H, estadio 44 (falanges osificadas); I-J, estadio 46 (I, esqueleto apendicular, los elementos del carpo se conservan cartilaginosos; J, detalle de la cintura escapular). Escala: 0,5 mm. Referencias: cl, clavícula; co, coracoides; cp, carpal-distal; ep, epicoracoides; es, escápula; eY, elemento Y; hu, húmero; me, metatarsal; pp, prepollex; pr, procoracoides; ra, radio; ral, radial; sp, supraescápula; ul, ulna; ulr, ulnar.
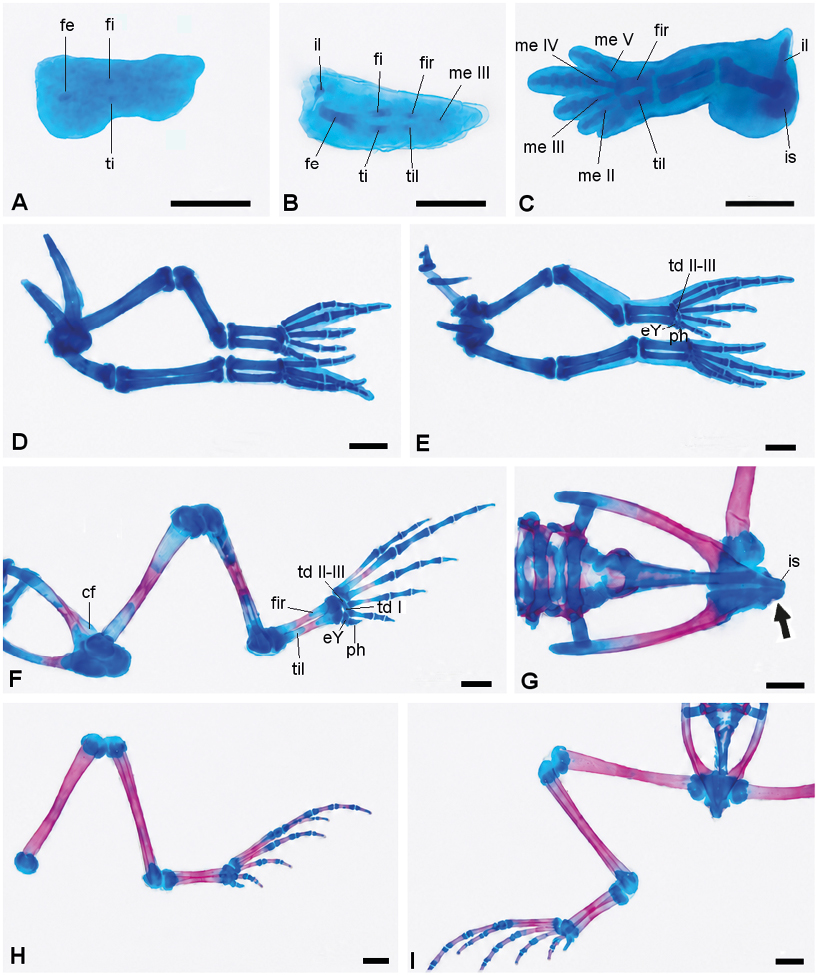
Figura 6. Secuencia de desarrollo de la extremidad posterior y cintura pélvica de Physalaemus biligonigerus (los elementos condrales están representados en azul y los elementos óseos, en rojo. A, estadio 32/33 (note los esbozos de elementos del estilopodio y zeugopodio); B, estadio 34/35 (se adicionan algunos componentes del autopodio y aparece un esbozo de la cintura pélvica); C, estadio 36/37 (se diferencian otros metatarsales, y la cintura pélvica adquiere forma de gancho); D, estadio 38/39 (aparición de los primeros centros de osificación en huesos largos, y de elementos del tarso); E, estadio 40/41 (avance de las osificaciones); F, estadio 42/43 (osificaciones en los metatarsales III-V, falanges cartilaginosas, tarso formado por tres piezas cartilaginosas, prehallux compuesto por dos elementos); G-H, estadio 44 (G, cintura pélvica, note el avance de la osificación del ilion, hacia el extremo distal, y las primeras osificaciones en el isquion-flecha-, H, extremidad posterior, note las falanges osificadas); I, estadio 46 (al final de la metamorfosis, el extremo anterior del ilion, la mayor parte del isquion, el pubis, y los tarsales continúan cartilaginosos). Escala: 0,5 mm. Referencias: cf, cresta femoral; eY, elemento Y; fa, falange; fe, fémur; fi, fíbula; fir, fibular; il, ilion; is, isquion; me, metatarsal; ph, prehallux; pu, pubis; td, tarsal distal; ti, tibia; til, tibial.
discusion
La comparación del desarrollo entre diferentes especies, únicamente sobre la base de estadios ontogenéticos, puede conducir a interpretaciones sesgadas, especialmente cuando algunos de los caracteres utilizados para definir dichos estadios se encuentran afectados por variaciones en el tiempo relativo de diferenciación. Desde una perspectiva evo-devo, resulta más apropiado comparar secuencias de desarrollo, ya que permiten evaluar el orden relativo de aparición de las estructuras, independientemente de marcadores cuyo momento de expresión puede variar entre linajes. Este enfoque reconoce que la ontogenia resulta de la interacción entre cambios en la forma, el tamaño y el tiempo, y que modificaciones en cualquiera de estas dimensiones pueden generar trayectorias de desarrollo divergentes entre especies (Haas, 1999; Fabrezi, 2012; Fabrezi et al., 2017b).
En Physalaemus biligonigerus, la osificación de los huesos craneales coincide con la aparición de las taenias tecti marginalis, transversalis y medialis, y con la aparición de centros de osificación en los huesos largos de las extremidades y en el ilion. La secuencia inicial de la osificación craneal sigue el patrón general descrito para la mayoría de los anuros metamórficos, donde los primeros elementos en osificarse corresponden al frontoparietal, al paraesfenoides y a los exoccipitales (Trueb y Alberch, 1985; Hanken y Hall, 1988; Moore y Townsend, 2003, Rocek 2003). Sin embargo, el orden temporal de osificación de estos elementos puede mostrar algunas variaciones entre las especies (Tabla I y II). Por ejemplo, en P. biligonigerus el primer componente en osificarse es el frontoparietal, en Pleurodema borelli, los frontoparietales y el paraesfenoides, mientras que en Leptodactylus macrosternum y L. latinasus, los exoccipitales (Perotti 2001, Gómez et al., 2017). En L. bufonius los exoccipitales se osifican luego de la metamorfosis (Vera y Ponssa, 2013). Mas allá de estas variaciones, en comparación con otros Leptodactylidae, en P. biligonigerus, estos eventos ocurren en etapas más avanzadas del desarrollo larval. Mientras que en L. macrosternum la mayoría de los huesos del cráneo son premetamórficos (excepto el neopalatino), en P. borelli se observa un estado intermedio, caracterizado por la presencia de cuatro elementos postmetamórficos. En contraste, Physalaemus biligonigerus presenta varios elementos en estado cartilaginoso hasta la metamorfosis, indicando un importante retardo en la osificación craneal (Tabla II). Entre los anuros, la presencia en estadios larvales de elementos del dermatocráneo, y únicamente osificaciones postmetamórficas del esfenoetmoides y opérculo, son considerados caracteres plesiomórficos. Especies consideradas basales dentro de los Anura, como Spea bombifrons, Rhinophrynus dorsalis y Xenopus laevis muestran patrones en los que la mayor parte del cráneo se encuentra osificado antes de la metamorfosis (Wiens, 1989; Trueb, 1985; Trueb y Hanken, 1992). Por el contrario, la condición en la que la osificación se completa luego de la metamorfosis, como en P. biligonigerus, está en concordancia con patrones heterocrónicos descritos para otros anuros (Hanken y Hall, 1988; Haas, 1999; Trueb, 1992).
Los cartílagos de la región oral están altamente especializados en renacuajos exotróficos (como los de P. biligonigerus), y constituyen adaptaciones asociadas a la alimentación en ecosistemas acuáticos (Wassersug y Hoff, 1982). Durante la metamorfosis, particularmente en los últimos estadios larvales, estas estructuras exhiben importantes remodelaciones, entre ellas la reabsorción de los suprarrostrales y fusión de los infrarrostrales con el cartílago de Meckel. Por otra parte, el esqueleto hiobranquial permanece cartilaginoso durante toda la etapa larval, y posteriormente es remodelado para originar el aparato hioideo del adulto. La persistencia de estas estructuras cartilaginosas hasta estadios larvales avanzados puede asociarse con la estrecha relación entre el desarrollo esquelético y los requerimientos funcionales de cada etapa onteogenética, acompañando así la transición desde una larva acuática hacia un individuo terrestre con hábitos alimentarios y demandas funcionales diferentes (Wassersug y Hoff, 1982; Hanken y Hall, 1988; Hall, 2012).
Con respecto al esqueleto postcraneal, el desarrollo de los miembros anteriores y de la cintura pectoral de P. biligonigerus es similar al de otras especies de hábitos terrestres, en tanto se originan a partir de las regiones adyacentes a la cavidad glenoidea (Haas, 1999). La diferenciación progresa desde la fosa glenoidea hacia los elementos más distales, y los primeros centros de osificación aparecen en el húmero y el radioulna, seguidos por los de la cintura. Las falanges y el cleitro son las últimas piezas en osificarse durante la fase larval, mientras que el epicoracoides, el procoracoides y los elementos carpales permanecen cartilaginosos hasta la metamorfosis. Este patrón, caracterizado por una progresión próximo-distal conservada, pero con variaciones en el tiempo de osificación de los elementos distales, es consistente con modelos evo-devo que destacan la organización modular del esqueleto apendicular y la mayor susceptibilidad de los módulos distales a cambios heterocrónicos durante la evolución del desarrollo (Haas, 1999; Fabrezi, 2011; Klingenberg, 2010; Fabrezi et al., 2017a). En este contexto, los miembros pueden considerarse módulos del desarrollo relativamente desacoplados de otros componentes corporales, permitiendo variaciones temporales y morfológicas asociadas a distintas trayectorias evolutivas.
La cintura pélvica presenta un patrón de desarrollo congruente con la hipótesis de un origen común de sus componentes (Manzano et al., 2013). Los primeros elementos en diferenciarse en P. biligonigerus son el ilion y el isquion, inicialmente observados como una unidad cartilaginosa continua, mientras que el pubis aparece posteriormente en continuidad con esta estructura. La osificación se inicia en el ilion y luego en el isquion, mientras que el pubis permanece cartilaginoso hacia el final de la metamorfosis. En comparación con Pleurodema borellii, algunos elementos de la cintura pélvica de P. biligonigerus muestran una diferenciación relativamente más tardía (Manzano et al., 2013).
En los miembros posteriores, las primeras osificaciones se observan en el estilopodio, el zeugopodio y los tarsales proximales (tibial y fibular), y hacia la metamorfosis los únicos elementos que aún se conservan cartilaginosos son los tarsales distales. La persistencia de estos elementos en estado cartilaginoso coincide con un patrón ampliamente distribuido entre los anuros, en el cual la mineralización y eventual osificación de los elementos modulares del carpo y tarso se prolonga durante la vida postmetamórfica (Fabrezi & Cruz, 2025).
Las diferencias observadas en el momento de diferenciación y osificación de algunos elementos pélvicos y apendiculares, junto con las registradas para ciertos componentes del cráneo, sugieren la existencia de variaciones heterocrónicas entre los distintos módulos esqueléticos. Este patrón podría constituir una importante fuente de variación ontogenética entre leptodactílidos y contribuir a la diversidad morfológica observada dentro del clado (Fabrezi, 2012; Fabrezi et al., 2017b).
En resumen, varias de las características observadas en P. biligonigerus coinciden con los patrones generales reconocidos en anuros terrestres (Haas, 1999), entre ellos el retraso relativo de la osificación de los miembros anteriores con respecto a los posteriores, la osificación temprana de las diáfisis de los huesos largos y persistencia en estado cartilaginoso de carpales y tarsales distales durante la fase larval. Esta condición ha sido documentada en diferentes géneros terrestres como Elachistocleis, Dermatonotus, Physalaemus, Leptodactylus, Trachycephalus, Scinax, Phyllomedusa, Lepidobatrachus, Ceratophrys y Chacophrys (Fabrezi, 2011). En contraste, en especies acuáticas como Pseudis platensis, las epífisis, los carpales y los tarsales completan su osificación y/o mineralización antes de la metamorfosis (Fabrezi y Goldberg, 2009). Estas diferencias entre taxones terrestres y acuáticas constituyen un patrón consistente de variación heterocrónica asociado al modo de vida, en el que la transición al ambiente terrestre se vincula con un retraso relativo en la maduración de los elementos distales del esqueleto apendicular (Haas, 1999; Fabrezi, 2011; Fabrezi et al., 2017a). Este patrón refleja la interacción entre restricciones del desarrollo y demandas funcionales asociadas a distintos modos locomotores, un proceso que habría contribuido a la diversificación morfológica de los anuros sin alterar la organización básica conservada del miembro.
Tabla I. Secuencia de osificación de elementos del cráneo durante la ontogenia postembrionaria de Physalaemus biligonigerus (este estudio) y otros géneros afines filogenéticamente, Pleurodema borelli (Perotti, 2011), Leptodactylus macrosternum (Perotti, 2011), L. bufonius (Vera y Ponssa, 2013) y L. latinasus (Gómez et al., 2011)
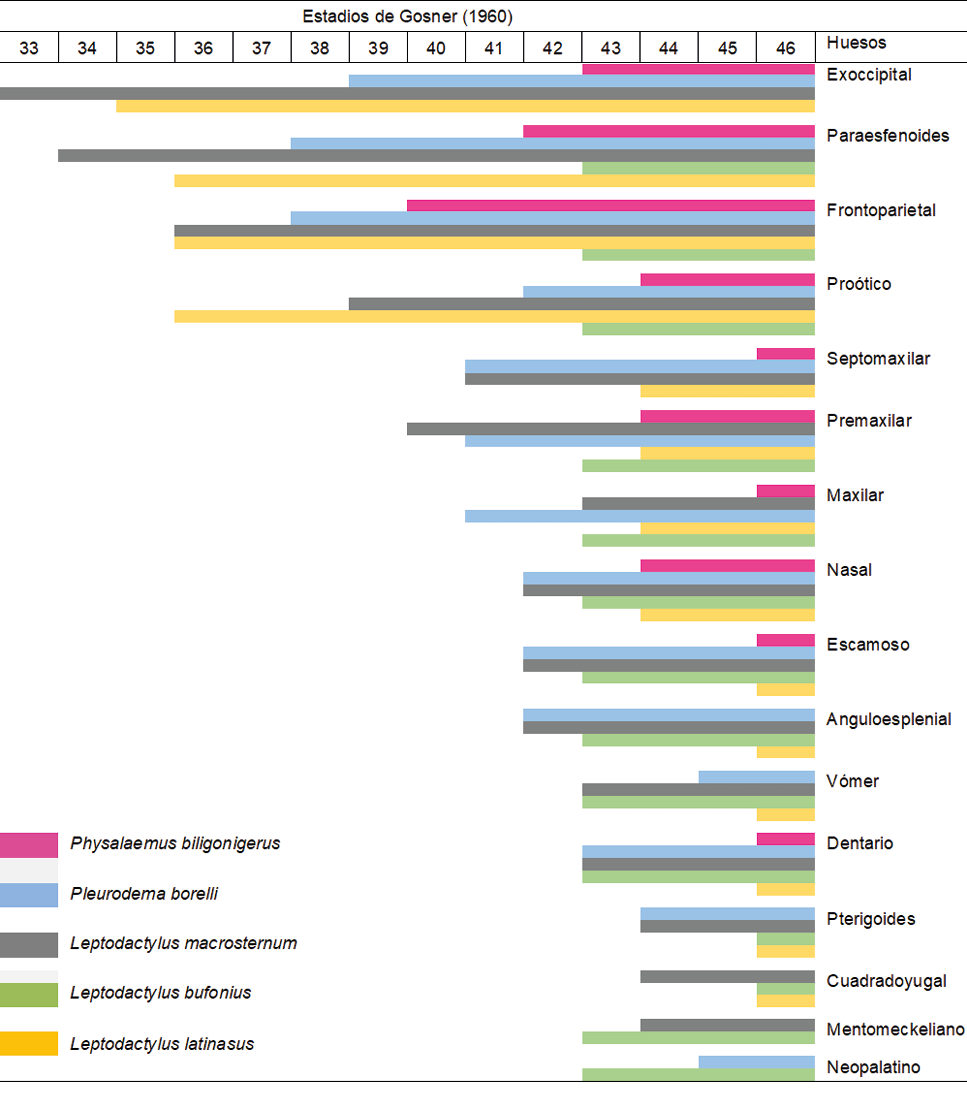
Tabla II. Secuencias de osificación (orden de aparición) de componentes del esqueleto craneal en Physalaemus biligonigerus y especies relacionadas filogenéticamente (dos o más elementos entre líneas indican huesos que se osifican en el mismo período de desarrollo).
|
P. biligonigerus |
|
P. borelli |
|
L. macrosternum |
|
L. bufonius |
|
L. latinasus |
|
Frontoparietal |
Frontoparietal |
Exoccipital |
Frontoparietal |
Exoccipital |
||||
|
Paraesfenoides |
Paraesfenoides |
Paraesfenoides |
Paraesfenoides |
Frontoparietal |
||||
|
Exoccipital |
Exoccipitales |
Frontoparietal |
Proótico |
Paraesfenoides |
||||
|
Proótico |
Septomaxila |
Proótico |
Premaxilar |
Proótico |
||||
|
Premaxilar |
Premaxilar |
Premaxilar |
Neopalatino |
Nasal |
||||
|
Nasal |
Maxilar |
Septomaxila |
Nasal |
Premaxilar |
||||
|
Escamoso |
Proótico |
Maxilar |
Dentario |
Maxilar |
||||
|
Dentario |
Nasal |
Nasal |
Anguloesplenial |
Septomaxilar |
||||
|
Maxilar |
Escamoso |
Escamoso |
Escamoso |
Dentario |
||||
|
Septomaxilar |
Angular |
Angular |
Mentomeckeliano |
Anguloesplenial |
||||
|
[Metamorfosis] |
Dentario |
Vómer |
Vómer |
Escamoso |
||||
|
Anguloesplenial |
Pterigoides |
Dentario |
Maxilar |
Pterigodes |
||||
|
Vómer |
Vómer |
Pterigoides |
Cuadradoyugal |
Vómer |
||||
|
Pterigoides |
Neopalatino |
Cuadrado yugal |
Pterigodes |
Cuadradoyugal |
||||
|
Cuadradoyugal |
[Metamorfosis] |
Mentomeckeliano |
[Metamorfosis] |
[Metamorfosis] |
||||
|
Columela |
Cuadrado yugal |
Columela |
Esfenoetmoides |
Mentomeckeliano |
||||
|
Mentomeckeliano |
Columela |
Hioides |
Exoccipital |
Palatino |
||||
|
Hioides |
Mentomeckeiano |
[Metamorfosis] |
Esfenoetmoides |
|||||
|
Neopalatino |
Hioides |
Neopalatino |
|
Columela |
||||
|
Este estudio |
Perotti, 2001 |
Perotti, 2001 |
Vera & Ponssa, 2013 |
Gómez et al., 2017 |
conclusiones
Physalaemus biligonigerus exhibe un patrón de condrificación y osificación que, aunque comparte rasgos generales con otros Leptodactylidae, presenta particularidades claras en el momento de aparición y en el grado de desarrollo de diversos elementos craneales y postcraneales. Las diferencias con especies cercanas ponen de manifiesto la diversidad de trayectorias ontogenéticas dentro de Leptodactylidae y resaltan la necesidad de comparar secuencias de diferenciación, un enfoque que permite evaluar el orden relativo de aparición de estructuras sin depender de estadios susceptibles a variación heterocrónica. Desde una perspectiva evo-devo, los desplazamientos temporales observados reflejan la acción de procesos heterocrónicos que modulan la maduración de distintos módulos esqueléticos y contribuyen a la diversidad morfológica dentro del clado. En conjunto, los patrones documentados para P. biligonigerus constituyen una base comparativa sólida para futuros estudios sobre heterocronía, evolución morfológica y relaciones filogenéticas en anuros.
Conflicto de intereses
Los autores declaran no tener conflicto de intereses.
Agradecimientos
A la Secretaría General de Ciencia y Técnica de la Universidad Nacional del Nordeste por el apoyo financiero. A la Subsecretaría de Recursos Naturales y Ecología (Dirección de Fauna y Parques, Ministerio de la Producción, provincia de Formosa) por los permisos otorgados oportunamente para la colección de las muestras. Este estudio se realizó en el marco del Trabajo Final de Graduación de OST para la obtención del título de Lic. en Ciencias Biológicas (FaCENA-UNNE).
Contribuciones de los autores
OSS llevó adelante la preparación y el análisis de las muestras, realizó las descripciones morfológicas y redactó la primera versión del manuscrito. VHZ colaboró en la interpretación de los resultados y revisó críticamente el manuscrito. Ambos autores contribuyeron de manera equivalente en la planificación general del trabajo y revisaron la versión final.
Arenas-Rodríguez, A., Rubiano Vargas, J. F., & Hoyos, J. M. (2018). Comparative description and ossification patterns of Dendropsophus labialis (Peters, 1863) and Scinax ruber (Laurenti, 1758) (Anura: Hylidae). PeerJ, 6, e4525.
Banbury, B., & Maglia, A. M. (2006). Skeletal development of the Mexican spadefoot, Spea multiplicata (Anura: Pelobatidae). Journal of Morphology, 267(7), 803-821.
de Sá, R. O. (1988). Chondrocranium and ossification sequence of Hyla lanciformis. Journal of Morphology, 195, 345-355.
de Sá, R. O., & Hill, S. (1998). Chondrocranial anatomy and skeletogenesis in Dendrobates auratus. Journal of Herpetology, 32 (2), 205–210.
de Sá, R. O., & Lavilla, E. O. (1996). Características de la osificación craneal en Phyllomedusa boliviana (Anura: Hylidae). Cuadernos de Herpetología, 9, 69-73.
de Sá, R. O., & Trueb, L. (1991). Osteology, skeletal development, and chondrocranial structure of Hamptophryne boliviana (Anura: Microhylidae). Journal of Morphology, 209, 311-330.
Fabrezi, M., Quinzio, S.I., Cruz, J. C., Chuliver Pereyra, M., Manzano, A., Abdala, V., Ponssa, M. L., Prieto, Y., & Goldberg, J. (2017b). Forma, tamaño y tiempo en la ontogenia de Anfibios y Reptiles. Cuadernos de Herpetología, 31(2), 103-126.
Fabrezi, M., Goldberg, J., & Pereyra, M. C. (2017a). Morphological variation in anuran limbs: Constraints and novelties. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution, 328(6), 546–574.
Fabrezi, M., & Barg, M. (2001). Patterns of carpal development among anuran amphibians. Journal of Morphology, 249, 210-220.
Fabrezi, M., & Goldberg, J. (2009). Heterochrony during skeletal development of Pseudis platensis (Anura: Hylidae) and the early offset of skeleton development and growth. Journal of Morphology, 270, 205-220.
Fabrezi, M., & Lavilla, E. O. (1993). Anatomía del chondrocráneo en larvas de tres especies de Telmatobius del grupo meridional (Anura: Leptodactylidae). Physis, 48, 39–46.
Fabrezi, M., & Vera, R. (1997). Caracterización morfológica de larvas de anuros del noroeste argentino. Cuadernos de Herpetología, 11, 37–49.
Fabrezi, M. (2011). Heterochrony in growth and development in anurans from the Chaco of South America. Evolutionary Biology, 38, 390-411.
Fabrezi, M. (2012). Heterocronía y variación morfológica en anuros. Cuadernos de Herpetología, 26, 29-47.
Fabrezi, M. (2001). A survey of prepollex and prehallux variation in anuran limbs. Zoological Journal of the Linnean Society, 131, 722–748.
Fabrezi, M., & Cruz, J. C. (2025). A comparative approach to the microstructure in the carpus and tarsus in anurans. Development, Growth & Differentiation, 67(2), 55–74.
Freemann, J., & de Sá, R. O. (1995). Chondrocranial morphology and ossification sequences of two species of Smilisca. University of Kansas Publications, Museum of Natural History, 17, 281–375.
Frost, D. R. (2025). Amphibian species of the world: An online reference (Version 5.5, January 31, 2025). American Museum of Natural History, New York, NY, USA. Retrieved from http://research.amnh.org/vz/herpetology/amphibia/
Goldberg, J., & Fabrezi, M. (2008). Development and variation of the anuran webbed feet (Amphibia, Anura). Zoological Journal of the Linnean Society, 152, 39–58.
Gómez, R. O., Regueira, E., O’Donohoe, M. E. A., & Hermida, G. N. (2017). Delayed osteogenesis and calcification in a large true toad with a comparative survey of the timing of skeletal ossification in anurans. Zoologischer Anzeiger, 267, 101–110.
Gosner, K. L. (1960). A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica, 16, 183–190.
Haas, A. (1999). Larval and metamorphic development in the fast-developing frog Pyxicephalus adspersus (Anura: Ranidae). Zoomorphology, 119, 23–35.
Haas, A. (2003). Phylogeny of frogs as inferred from primarily larval characters (Amphibia: Anura). Cladistics, 19, 23–89.
Hall, J. A., & Larsen, J. H., Jr. (1998). Postembryonic ontogeny of the spadefoot toad, Scaphiopus intermontanus (Anura: Pelobatidae): Skeletal morphology. Journal of Morphology, 238(2), 179–244.
Hall, B. K. (2012). Evolutionary developmental biology (Evo-Devo): Past, present, and future. Evolution: Education and Outreach, 5(2), 184–193.
Hanken, J., & Hall, B. K. (1988). Skull development during anuran metamorphosis: I. Early development of the first three bones to form—the exoccipital, parasphenoid, and frontoparietal. Journal of Morphology, 195(3), 247–256.
Hanken, J., & Hall, B. K. (1988). Skull development during anuran metamorphosis. I. Early development of the first three bones to form: The exoccipital, the parasphenoid, and the frontoparietal. Journal of Morphology, 195, 247–256.
Kerney, R., Meegaskumbura, M., Manamendra-Arachchi, K., & Hanken, J. (2007). Cranial ontogeny in Philautus silus (Anura: Ranidae: Rhacophorinae) reveals few similarities with other direct-developing anurans. Journal of Morphology, 268, 715–725. [URL]>
Klingenberg, C. P. (2010). Evolution and development of shape: Integrating quantitative approaches. Nature Reviews Genetics, 11(9), 623–635.
Kovalenko, E. E., & Kruzhkova, I. U. (2013). Individual variation in the development of the common toad, Bufo bufo (Anura, Bufonidae): 2. Diagnostic characters of the axial skeleton. Ontogenez, 44(4), 249–264.
Kuratani, S. (2009). Modularity, comparative embryology and evo-devo: Developmental dissection of evolving body plans. Developmental Biology, 332(1), 61–69.
Larson, P., & de Sá, R. O. (1998). Chondrocranial morphology of Leptodactylus larvae (Leptodactylidae: Leptodactylinae): Its utility in phylogenetic reconstruction. Journal of Morphology, 238, 287–305.
Lavilla, E. O., & Fabrezi, M. (1987). Anatomía de larvas de Hyla pulchella andina (Anura: Hylidae). Physis, 45, 77–82.
Lourenço, L.B., Targueta, C.P., Baldo, D., Nascimento, J., Garcia, P.C., Andrade, G.V., Haddad, C.F., & Recco‐Pimentel, S.M. (2015). Phylogeny of frogs from the genus Physalaemus (Anura, Leptodactylidae) inferred from mitochondrial and nuclear gene sequences. Molecular phylogenetics and evolution, 92, 204–216.
Maglia, A. M., Pugener, L. A., & Trueb, L. (2001). Comparative development of anurans: Using phylogeny to understand ontogeny. American Zoologist, 41, 538–551.
Maglia, A. M., & Pugener, L. A. (1998). Skeletal development and adult osteology of Bombina orientalis (Anura: Bombinatoridae). Herpetologica, 54, 344–363.
Manzano, A., Abdala, V., Ponssa, M.L., & Soliz, M. (2013). Ontogeny and tissue differentiation of the pelvic girdle and hind limbs of anurans. Acta Zoologica (Stockholm) 94, 420–436.
Moore, M. K., & Townsend, V. (2003). Intraspecific variation in cranial ossification in the tailed frog, Ascaphus truei. Journal of Herpetology, 37, 714–717.
Müller, G. B. (2007). Evo–devo: Extending the evolutionary synthesis. Nature Reviews Genetics, 8(12), 943–949.
Perotti, M. G. (2001). Skeletal development of Leptodactylus chaquensis (Anura: Leptodactylidae). Herpetologica, 57, 318–335.
Púgener, L. A., & Maglia, A. M. (1997). Osteology and skeletal development of Discoglossus sardus (Anura: Discoglossidae). Journal of Morphology, 233, 267–286.
Rôcek, Z., Böttcher, R., & Wassersug, R. J. (2006). Gigantism in the tadpoles of the Neogene frog Paleobatrachus. Paleobiology, 32(4), 666–675.
Senevirathne, G., Kerney, R., & Meegaskumbura, M. (2017). Comparative postembryonic skeletal ontogeny in two sister lineages of Old World tree frogs (Rhacophoridae: Taruga, Polypedates). PLoS ONE, 12(1), e0167939.
Sheil, C. A., & Alamillo, H. (2005). Osteology and skeletal development of Phyllomedusa vaillanti (Anura: Hylidae: Phyllomedusinae) and a comparison of this arboreal species with a terrestrial member of the genus. Journal of Morphology, 265(3), 343–368.
Sheil, C. A. (1999). Osteology and skeletal development of Pyxicephalus adspersus (Anura: Ranidae: Raninae). Journal of Morphology, 240, 49–75.
Smith, K. K. (2003). Time’s arrow: Heterochrony and the evolution of development. International Journal of Developmental Biology, 47, 613–621.
Stokely, P. S., & List, J. C. (1954). The progress of ossification in the skull of the cricketfrog Pseudacris nigrita triseriata. Copeia, 1954, 211–217.
Talavera, R. R. (1985). Notas sobre la ontogenia de las especializaciones locomotoras del tarso de Pelodytes punctatus (Anura). Butlletí de la Societat Catalana d’Ictiologia i Herpetologia, 10, 27–31.
Trueb, L., Púgener, L. A., & Maglia, A. M. (2000), Ontogeny of the bizarre: an osteological description of Pipa pipa (Anura: Pipidae), with an account of skeletal development in the species. Journal of Morphology, 243: 75-104.
Trueb, L., & Alberch, P. (1985). Miniaturization and the anuran skull: A case study of heterochrony. In H. R. Duncker y G. Fleischer (Eds.), Vertebrate Morphology. Fortschritte der Zoologie (Vol. 30, pp. 113–121). Stuttgart, NY: Gustav Fischer Verlag.
Trueb, L., & Hanken, J. (1992). Skeletal development in Xenopus laevis (Anura: Pipidae). Journal of Morphology, 214(1), 1–41.
Trueb, L. (1985). A summary of osteocranial development in anurans with notes on the sequence of cranial ossification in Rhinophrynus dorsalis (Anura: Pipoidea: Rhynophrynidae). South African Journal of Science, 81, 181–185.
Trueb, L. (1992). Patterns of cranial diversity among the Lissamphibia. In J. Hanken y B. K. Hall (Eds.), The vertebrate skull: Patterns of structural and systematic diversity. Vol. I: Structural and systematic diversity (pp. 255–343). University of Chicago Press.
Vera Candioti, M. F., Úbeda, C., & Lavilla, E. O. (2005). Morphology and metamorphosis of Eupsophus calcaratus tadpoles (Anura: Leptodactylidae). Journal of Morphology, 264, 161–177.
Vera, M. C., & Ponssa, M. L. (2013). Skeletogenesis in anurans: Cranial and postcranial development in metamorphic and postmetamorphic stages of Leptodactylus bufonius (Anura: Leptodactylidae). Acta Zoologica (Stockholm), 95, 44–62.
Wassersug, R. J., & Hoff, K. (1982). Developmental changes in the orientation of the anuran jaw suspension: A preliminary exploration into the evolution of anuran metamorphosis. Evolutionary Biology, 15, 223–246.
Wassersug, R. L. (1976). A procedure for differential staining of cartilage and bone in whole formalin fixed vertebrates. Stain Technology, 51, 131–134.
Wiens, J. J. (1989). Ontogeny of the skeleton of Spea bombifrons (Anura: Pelobatidae). Journal of Morphology, 202, 29-51.
Wild, E. R. (1999). Description of the chondrocranium and osteogenesis of the Chacoan burrowing frog (Anura: Leptodactylidae). Journal of Morphology, 242(3), 229–246.
Wild, E. R. (1997). Description of the adult skeleton and developmental osteology of the hyperossified horned frog, Ceratophrys cornuta (Anura: Leptodactylidae). Journal of Morphology, 232(2), 169–206.
Yıldırım, E., & Kaya, U. (2016). Development of the limb skeleton of Bufotes variabilis (Anura: Bufonidae) and comparison with other bufonids. Russian Journal of Herpetology, 23, 215–223.