Articulo original
Original article
36 (1) | Ene - Jun, 2026
Facultad de Ciencias Exactas y Naturales y Agrimensura (UNNE)
Open Access: https://revistas.unne.edu.ar/index.php/fce
E-mail: revistafacena@exa.unne.edu.ar
Osmotic stress tolerance in two Neotropical anurans: effects of salinity on larval growth and development
Tolerancia al estrés osmótico en dos anuros Neotropicales: efectos de la salinidad en el crecimiento y desarrollo larvario
Marangoni, Federico * ¹ & Gómez, Valeria I. ²
1 Grupo de Investigación en Anfibios y Reptiles de la Facultad de Ciencias Exactas y Naturales y Agrimensura (FaCENA, UNNE), CONICET. Corrientes, Corrientes, Argentina.
2 Laboratorio de Ecología y Diversidad de Anfibios y Reptiles del NEA, Centro de Ecología Aplicada del Litoral (LEDAR), UNNE, CONICET. Corrientes, Corrientes, Argentina.
* Autor de correspodencia: fedemarangoni@gmail.com
Recibido/Received: 17 de abril, 2026 | Aceptado/Acepted: 25 de junio, 2026 | Publicado/Published: 08 de julio, 2026
Como citar este artículo: Marangoni, F. & Gómez, V. I. (2026). Osmotic stress tolerance in two Neotropical anurans: effects of salinity on larval growth and development. Revista FACENA 36(1), 140-161. Doi: https://doi.org/10.30972/fac.3619271
Resumen: Los ecosistemas de agua dulce están cada vez más amenazados por la salinización derivada de actividades humanas y el cambio climático, con importantes consecuencias para la biodiversidad acuática. Los anfibios son particularmente vulnerables debido a su piel permeable y a sus etapas tempranas de vida dependientes del medio acuático, aunque las respuestas de especies neotropicales aún son poco conocidas. Evaluamos los efectos letales y subletales de la salinidad en embriones y larvas de Rhinella dorbignyi y Leptodactylus macrosternum. Se realizaron experimentos de dosis–respuesta en laboratorio utilizando gradientes de NaCl (0–12 g/l) para estimar LC₅₀, y ensayos subletales (0–8 g/l) para evaluar supervivencia, crecimiento y desarrollo. Los individuos provinieron de poblaciones sintópicas y fueron criados en condiciones controladas. La salinidad incrementó la mortalidad en ambas especies en etapas embrionarias y larvales, con valores de LC₅₀ entre 6–9,7 g/l en R. dorbignyi y 7,1–8,8 g/l en L. macrosternum. Se observó una disminución de la tolerancia dependiente del tiempo más marcada en R. dorbignyi. En los ensayos subletales, la supervivencia disminuyó con el aumento de salinidad, especialmente en R. dorbignyi, mientras que los efectos en L. macrosternum fueron más débiles. La masa corporal, la tasa de crecimiento y de desarrollo se redujeron significativamente en R. dorbignyi, mientras que L. macrosternum mostró cambios limitados o no significativos. En conjunto, la salinidad actuó como una fuerte presión selectiva sobre el desempeño temprano, con claras diferencias interespecíficas en sensibilidad. Estos resultados indican que incluso incrementos moderados de salinidad en charcas temporales pueden reducir el reclutamiento, y que la duración de la exposición es un factor clave. Se destaca la necesidad de integrar respuestas letales y subletales en ecotoxicología, incorporando múltiples poblaciones por especie y un mayor muestreo taxonómico, incluyendo variación intraespecífica, para distinguir plasticidad de adaptación local y mejorar las predicciones bajo escenarios de cambio ambiental.
Palabras claves: Anfibios; Rhinella dorbignyi; Leptodactylus macrosternum; Salinidad; Estrés; Argentina.
Abstract: Freshwater ecosystems are increasingly threatened by salinization driven by human activities and climate change, with major consequences for aquatic biodiversity. Amphibians are particularly vulnerable due to permeable skin and aquatic-dependent early life stages, yet Neotropical responses remain poorly understood. We evaluated lethal and sublethal effects of salinity on embryos and larvae of Rhinella dorbignyi and Leptodactylus macrosternum. Laboratory dose–response experiments using NaCl gradients (0–12 g/l) estimated LC₅₀, and larval assays under sublethal salinity (0–8 g/l) assessed survival, growth, and development. Individuals from syntopic populations were reared under controlled conditions. Salinity increased mortality in both species at embryonic and larval stages, with LC₅₀ values ranging from 6–9.7 g/l in R. dorbignyi and 7.1–8.8 g/l in L. macrosternum. A time-dependent decline in tolerance was more evident in R. dorbignyi. In sublethal assays, survival decreased with increasing salinity, particularly in R. dorbignyi, while effects in L. macrosternum were weaker. Larval body mass, growth, and developmental rate were significantly reduced under salinity exposure in R. dorbignyi, whereas L. macrosternum showed limited or non-significant changes in growth and development. Overall, salinity acted as a strong selective pressure affecting early-life performance, with clear interspecific differences in sensitivity. These results indicate that even moderate salinity increases in temporary ponds can reduce recruitment and that exposure duration is a key driver of impact. We emphasize integrating lethal and sublethal endpoints in ecotoxicological assessments and highlight that future studies should incorporate multiple populations per species and broader taxonomic sampling, including tests of intraspecific variation in tolerance, to disentangle plasticity from local adaptation and improve predictions under environmental change.
Keywords: Amphibians; Rhinella dorbignyi; Leptodactylus macrosternum; Salinity; Stress; Argentina.
INTRODUCTION
Freshwater ecosystems are among the most threatened environments worldwide, facing increasing pressures from anthropogenic activities and climate change (Dudgeon et al., 2006; Vörösmarty et al., 2010; Calhoun et al., 2017). Among these stressors, the salinization of inland waters has emerged as a significant and growing ecological concern, driven by factors such as deforestation, agricultural runoff, urbanization, and rising evaporation rates (Kaushal et al., 2005; Cañedo-Argüelles et al., 2013; Herbert et al., 2015). This process can profoundly alter the physicochemical properties of aquatic habitats, affecting species distribution, community structure, and ecosystem functioning (Biggs et al., 2017).
Amphibians are particularly vulnerable to changes in environmental salinity due to their highly permeable skin and complex life cycles that depend on aquatic environments for reproduction and early development (Hopkins & Brodie, 2015; Relyea et al., 2024). Embryonic and larval stages are especially sensitive to osmotic stress, which can disrupt ion balance, increase energetic costs, and ultimately impair survival, growth, and development (Balinsky, 1981; Boutilier et al., 1992; Gomez-Mestre & Tejedo, 2003; Stănescu et al., 2017). Experimental studies have shown that increased salinity can reduce larval survival, delay growth and development, decrease activity, and affect morphological traits in a wide range of amphibian species (Gomez-Mestre & Tejedo, 2003; Gomez-Mestre et al., 2004; Sanzo & Hecnar, 2006; Chinathamby et al., 2006; Karraker et al., 2008; Wu & Kam, 2009). Moreover, salinity may influence the coexistence of sympatric species by differentially affecting their reproductive success and fitness (Stănescu et al., 2017).
Life-history traits such as growth rate, developmental timing, and size at metamorphosis are key determinants of fitness in amphibians (Semlitsch et al., 1988; Wells, 2007). Environmental stressors like salinity can alter these traits, potentially leading to long-term consequences for individual performance and population dynamics. For example, slower growth or delayed metamorphosis may increase exposure to predation and desiccation risks, while reduced body size at metamorphosis can negatively impact survival and reproductive success (Székely et al., 2020; Burraco et al., 2022).
Despite these generally negative effects, evidence suggests that amphibians may exhibit considerable variation in their responses to salinity stress. Such variation may arise from the evolution of tolerance to increasing salinization, phenotypic plasticity, or local adaptation (Gomez-Mestre & Tejedo, 2003; Relyea et al., 2023, 2024). The studies have shown that exposure to saline environments can drive rapid shifts in tolerance, with certain anuran amphibian species displaying variable and adaptive responses to salt exposure across populations and among different life stages (Albecker & McCoy, 2017; Relyea et al., 2023, 2024). These findings underscore the complexity of organismal responses to salinization and the importance of considering both ecological and evolutionary perspectives.
In addition, the effects of salinity are often modulated by interactions with other environmental stressors. For instance, salinity stress has been shown to reduce thermal tolerance in amphibian larvae, suggesting potential synergistic effects under climate change scenarios (Sanabria et al., 2018; Chuang et al., 2022). Similarly, elevated salinity levels can exacerbate disease dynamics, increasing the severity of pathogen outbreaks such as ranavirus infections in amphibian populations (Hall et al., 2020).
Despite growing interest in the ecological and evolutionary effects of salinization, significant knowledge gaps remain. In particular, fewer studies have simultaneously evaluated lethal and sublethal responses across multiple taxa (Hintz & Relyea, 2019). Additionally, information from Neotropical amphibians remains scarce, limiting our ability to generalize patterns and predict responses in these highly diverse and ecologically important assemblages. Addressing these gaps is critical, especially in regions where freshwater systems are increasingly exposed to salinization but remain understudied.
In this study, we evaluate the effects of salinity on early life stages of two Neotropical anurans, Rhinella dorbignyi and Leptodactylus macrosternum, adopting an eco-physiological approach. Specifically, we assess (1) the relationship between salinity and mortality through dose–response experiments, and (2) the effects of sublethal salinity exposure on larval growth and developmental rates. Given the known sensitivity of early life stages and the energetic costs associated with osmoregulation, we predict that increasing salinity will reduce survival and negatively affect growth and development in both species. Furthermore, we expect interspecific differences in tolerance, reflecting variation in their ecological and physiological characteristics.
MATERIALS AND METHODS
Sampling site
Fieldwork was carried out in a peri-urban area surrounding the airport of the city of Corrientes (27°27’03.2” S 58°46’26.3” W), where Rhinella dorbignyi and Leptodactylus macrosternum occur in syntopy. This area is characterized by the presence of numerous temporary ponds that fill with rainfall at the beginning of spring and may persist for 15–30 days (Fig. 1a). It is a natural area in which the original vegetation consisted of Schinopsis balansae (“quebracho”) forest, currently highly degraded due to cattle, pig, and sheep grazing. In addition, the site is frequently subjected to intentional burning by local inhabitants to promote pasture renewal for livestock. Mean annual temperature is 21.5°C and the mean annual precipitation is 1500 mm, without a pronounced dry season, although periods of rain shortages occur every 4-6 years (Carnevali, 1994).On 3 and 24 August 2025, we sampled one recently laid clutch of Rhinella dorbignyi and Leptodactylus macrosternum, respectively, at a comparable developmental stage (Gosner stage 10; Gosner, 1960). Approximately 10% of the total clutch of R. dorbignyi was carefully removed, whereas the entire foam nest of L. macrosternum was collected and transported to the LEDAR laboratory at the CECOAL-UNNE/CONICET. Embryos were maintained in plastic trays containing dechlorinated tap water. Room temperature was kept approximately constant (23–25 °C), and the photoperiod followed a natural cycle (L10:D14).
Lethal concentration 50% experiments in Embryonic and Early Larval Stages
We used the median lethal concentration (LC₅₀), defined as the concentration causing 50% mortality (Sparks, 2000), to assess the embryonic and larval salinity tolerance limits of the two anuran species studied. Accordingly, we conducted two independent classical acute toxicity tests for each species: one at the embryonic stage and another at the larval stage. We followed the experimental design and salinity gradient described by Gómez-Mestre and Tejedo (2003), with slight modifications. The salinity gradient was established using separate plastic ice cube trays (Fig. 1B). For both embryonic and larval tests, five salinity levels were used: a freshwater control and 6, 8, 10, and 12 g/l total dissolved solids (commercial salt, NaCl). We randomly assigned 36 embryos and larvae to each treatment (Fig. 1B). Mortality was recorded at 24, 48, and 72 h after exposure to the salinity gradient. Embryonic toxicity tests were initiated on the same day the clutches were collected (3 and 24 August 2025 for R. dorbignyi and L. macrosternum, respectively), when embryos of both species were at development stage 10 (Gosner, 1960). Larval tests were initiated once individuals of both species reached development stage 26 (Gosner, 1960), on 18 August and 2 September for R. dorbignyi and L. macrosternum, respectively. Data were analyzed by nonlinear regression between mortality and salinity gradient (treatments) (Sokal and Rohlf, 1995), fitting an exponential model to the observations. Mortality was calculated as the ratio of dead embryos to the initial number of embryos, and LC₅₀ values were derived from the regression equation using Excel. Analyses were performed using the Statistica software package (StatSoft 2007). Nonlinear regressions and LC₅₀ calculations were performed using Microsoft Excel (Microsoft Corporation, Redmond, WA, USA). Differences in mortality among salinity treatments were evaluated using a chi-square (χ²) test of independence, based on the number of surviving and dead individuals per treatment. Analyses were performed in Microsoft Excel (CHISQ.TEST), with significance set at α = 0.05.
Larval tolerance experiments
Experiments were designed to test the tolerance of both species to sublethal water salinity during the larval phase (from Gosner stage 26). Four salinity levels were used: a freshwater control and 2, 4, and 6 g/l, prepared as described for the previous experiment. Larvae were obtained from the same clutches used in the previous experiment. For each treatment, 36 tadpoles per species were randomly selected and assigned to each treatment. Tadpoles were individually placed in 400 cm³ plastic cups containing dechlorinated tap water and randomly assigned to shelves of a metal rack (Fig. 1C and D). The experiment was initiated once individuals of both species reached development stage 26 (Gosner, 1960), on 18 August and 2 September for R. dorbignyi and L. macrosternum, respectively. Tadpoles were fed 0.1 g of commercial fish food (Sera Micron, Sera GmbH, Germany) daily. We deployed two temperature data loggers (HOBO Pendant): one in the laboratory to record air temperature and the other inside a plastic cup containing dechlorinated tap water to record water temperature during the course of the experiments. Temperature was recorded every 1 h. Body mass, developmental stage (Gosner, 1960) and mortality, were recorded at 10 and 20 days after exposure to the salinity gradient. Body mass was measured using a precision balance (±0.001 g; OHAUS Adventurer™, OHAUS. Corporation, USA). Developmental stages were determined using a stereomicroscope (Leica EZ4, Leica Microsystems, Wetzlar, Germany). Larval mortality was analyzed using a chi-square (χ²) test of independence, following the same procedure described above for embryonic mortality. Growth and developmental rate were defined as (initial mass or stage – mass or stage at 10 or 20 days) / 10 or 20 (depending on the exposure time). All variables were log-transformed in order to meet the normality and homoscedasticity assumptions. We used multi- and univariate analyses of variance to test for differences in body mass, growth and developmental rate, between treatments and species. Means were compared using a post hoc Scheffe´ multiple comparison test, at a=0.05. All statistical analyses were analyzed using the Statistica 6.0 statistical package (StatSoft, 2007).

Figure 1. Sampling site (A), design of toxicity tests in the embryonic phase (B) and assessment of salinity effects on larval growth and development rate (C and D).
results
Lethal concentration 50% experiments in Embryonic and Early Larval Stages
Figure 2 graphically shows the LC₅₀ values derived from the regression equations obtained in the embryonic and larval LC₅₀ experiments. After 24, 48, and 72 h of exposure, embryonic LC₅₀ values in Rhinella dorbignyi ranged from 6 to 9.7 g/l, whereas larval LC₅₀ values ranged from 8.21 to 8.76 g/l. In Leptodactylus macrosternum, LC₅₀ values ranged from 7.1 to 8.1 g/l for embryos and from 8.10 to 8.8 g/l for larvae. Water salinity had a clear effect on embryonic and larval mortality (Table 1, Fig. 2). In both phases of the two species, a positive relationship between salinity and mortality was observed (Fig. 2). Regarding the relationship between LC₅₀ values and exposure time, a negative trend was observed in embryos and larvae of R. dorbignyi. In other words, 50% mortality was reached at lower salinity concentrations as exposure time increased. However, this pattern was not observed in L. macrosternum.
Table 1. Results of chi-square (χ²) tests evaluating differences in mortality among salinity treatments for both studied species. Values correspond to χ² statistics, degrees of freedom (df), and associated significance levels. All tests were based on the number of surviving and dead individuals per treatment, with significance set at α = 0.05.
|
Rhinella dorbignyi |
Leptodactylus macrosternum |
|
|
LC50 Embryonic |
||
|
24 h |
χ² = 96.10, df = 4, P < 0.001 |
χ² = 144.42, df = 4, P < 0.001 |
|
48 h |
χ² = 85.06, df = 4, P < 0.001 |
χ² = 165.43, df = 4, P < 0.001 |
|
72 h |
χ² = 134.88, df = 4, P < 0.001 |
χ² = 159.60, df = 4, P < 0.001 |
|
LC50 Larval |
||
|
24 h |
χ² = 166.52, df = 4, P < 0.001 |
χ² = 172.64, df = 4, P < 0.001 |
|
48 h |
χ² = 150.01, df = 4, P < 0.001 |
χ² = 121.87, df = 4, P < 0.001 |
|
72 h |
χ² = 131.94, df = 4, P < 0.001 |
χ² = 102.27, df = 4, P < 0.001 |
|
Larval Experiments |
||
|
10 days |
χ² = 104.72, df = 3, P < 0.001 |
χ² = 3.04, df = 3, P = 0.385 |
|
20 days |
χ² = 119.54, df = 3, P < 0.001 |
χ² = 2.94, df = 3, P = 0.401 |
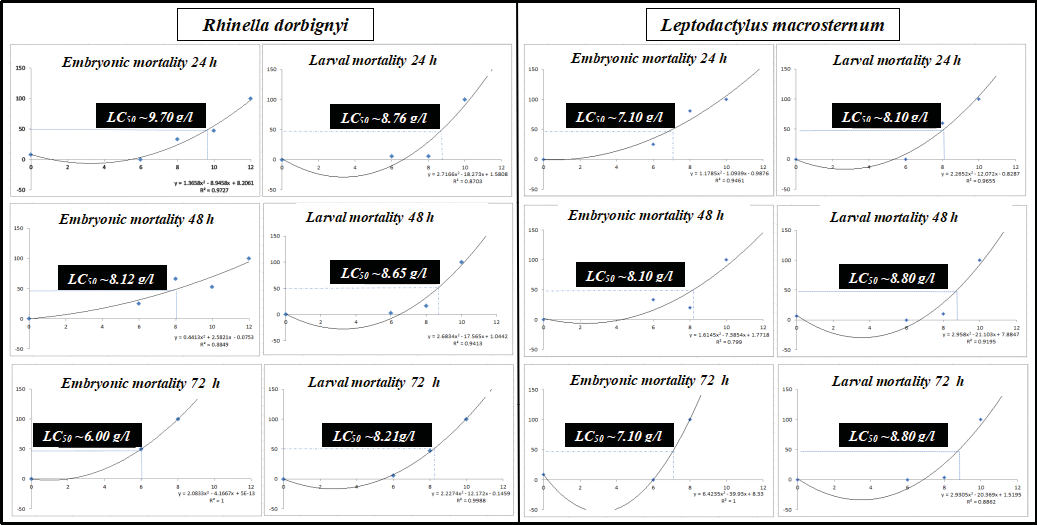
Figure 2. Results of the LC50 experiment after the first 24, 48 and 72 hours in Rhinella dorbignyi and Leptodactylus macrosternum. The x-axis represents the water salinity gradient ranging from freshwater to 12 g/l. The y-axis represents mortality (as a proportion of the initial number of embryos per replicate) of the embryos at each of the stress doses. A polinomic model was fit to each of the populations. The LC50 value for each population is graphically approximated by the dotted line depicted in each plot.
Larval tolerance experiments
Water salinity had a significant effect on larval survival in R. dorbignyi, with increasing salinity levels leading to higher mortality relative to the freshwater control group (Table 1). This effect intensified with increasing exposure time (Fig. 3). However, the magnitude of these responses differed between species, with comparatively weaker effects in L. macrosternum (Fig. 3), where no significant differences among treatments were observed (Table 1). In R. dorbignyi, survival was markedly reduced at 6 g/l after 10 days of exposure and declined further by day 20, both at this concentration and at 4 g/l. In contrast, although mortality in L. macrosternum increased over time, differences among salinity treatments and the control group remained relatively small (Fig. 3).
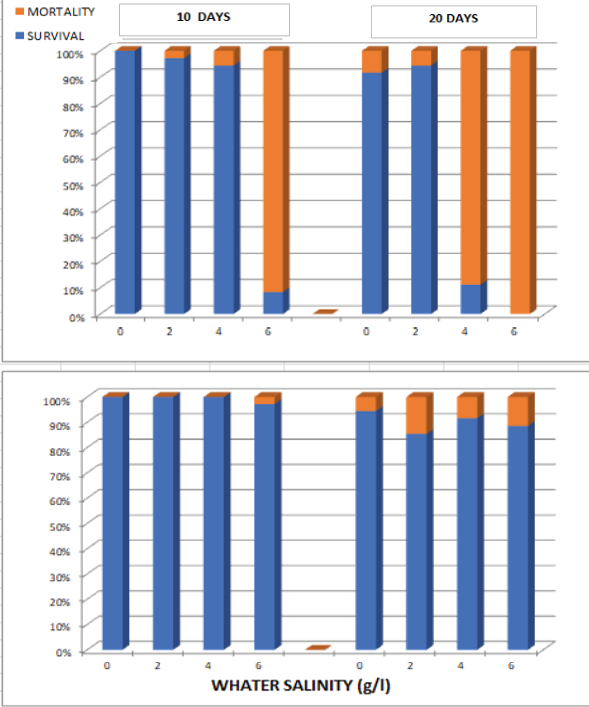
Figure 3. Mortality/Survival at 10 and 20 days of exposure to different sublethal concentrations during the larval stage of Rhinella dorbignyi (top) and Leptodactylus macrosternum (bottom).
Effects of salinity on larva body mass
The descriptive statistics of larval body mass at 10 and 20 days of exposure to different sublethal concentrations in Rhinella dorbignyi and Leptodactylus macrosternum are presented in Table 2. Water salinity negatively affected larval body mass, with marked differences between species (Fig. 4). Although body mass increased from the beginning of the experiment to day 20 in both species (Fig. 4), body mass was consistently lower across all salinity treatments at the end of the experiments in R. dorbignyi (differences between treatments ANOVA: F2, 68 = 17.663, P < 0.0001; Table 2). In contrast, in L. macrosternum, although differences between treatments were observed (ANOVA: F3, 96 = 16.317, P < 0.0001), the body mass of the larvae from control group did not differ significantly from those exposure to the highest (6 g/l) and lowest (2 g/l) salinity treatments (Table 2). The percentage of body mass attained after 10 days of the experiment, relative to total mass, is shown in Fig. 4.
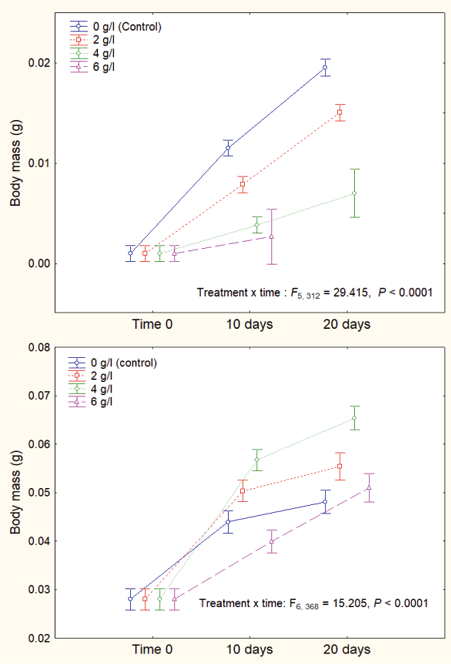
Figure 4. Body mass recorded at 10 and 20 days of exposure to different sublethal concentrations in Rhinella dorbignyi (top) and Leptodactylus macrosternum (bottom). The percentage of body mass attained after 10 days of the experiment, relative to total mass, is shown.
Table 2. Descriptive statistics of larval growth at 10 and 20 days of exposure to different sublethal concentrations in Rhinella dorbignyi and Leptodactylus macrosternum. SE = standard error. CI = confidence interval. Different letters in bold mean significant differences using a Scheffe´ post hoc multiple comparison test at α = 0.05.
|
Time elapsed |
Larval body mass (g) |
|||||
|
Mean (g) |
SE |
CI -95% |
CI +95% |
N |
||
|
Rhinella dorbignyi |
||||||
|
0 g/l (Control) |
10 days |
0.0115 |
0.0004 |
0.0107 |
0.0123 |
36 |
|
0 g/l (Control) |
20 days |
0.0195(a) |
0.0004 |
0.0187 |
0.00203 |
33 |
|
2 g/l |
10 days |
0.0079 |
0.0004 |
0.0071 |
0.0087 |
35 |
|
2 g/l |
20 days |
0.0151(b) |
0.0004 |
0.0142 |
0.0159 |
34 |
|
4 g/l |
10 days |
0.0038 |
0.0004 |
0.0030 |
0.0046 |
34 |
|
4 g/l |
20 days |
0.0070(c) |
0.0012 |
0.0046 |
0.0094 |
4 |
|
6 g/l |
10 days |
0.0027 |
0.0014 |
-0.0001 |
0.0054 |
3 |
|
6 g/l |
20 days |
---- |
---- |
---- |
---- |
---- |
|
Leptodactylus macrosternum |
||||||
|
0 g/l (Control) |
10 days |
0.0439 |
0.0012 |
0.0416 |
0.0462 |
33 |
|
0 g/l (Control) |
20 days |
0.0481(a) |
0.0012 |
0.0456 |
0.0505 |
29 |
|
2 g/l |
10 days |
0.0503 |
0.0011 |
0.0481 |
0.0526 |
35 |
|
2 g/l |
20 days |
0.0554(a) |
0.0014 |
0.0526 |
0.0582 |
22 |
|
4 g/l |
10 days |
0.0567 |
0.0011 |
0.0545 |
0.0589 |
36 |
|
4 g/l |
20 days |
0.0653(b) |
0.0012 |
0.0629 |
0.0678 |
29 |
|
6 g/l |
10 days |
0.0399 |
0.0012 |
0.0375 |
0.0422 |
32 |
|
6 g/l |
20 days |
0.0510(a) |
0.0015 |
0.0480 |
0.0540 |
20 |
Effects of salinity on larval growth and development rates
Water salinity (treatments) showed significant differences regarding growth and development rates in both species (MANOVA: R. dorbignyi: Wilk’s λ = 0.803, F6, 750 = 14.467, P < 0.001; L. macrosternum: Wilk’s λ = 0.647, F6, 636 = 25.802, P < 0.001). Univariate ANOVA F-values for growth and development of R. dorbignyi and L. macrosternum are summarized in Table 3. Lower growth rates were observed under salinity treatments in R. dorbignyi; however, no significant differences were found (post-hoc Scheffé: P = 0.214) between the 6 g/l treatment and the control group in L. macrosternum (Fig. 5). Lower development rate also was observed under salinity treatments in R. dorbignyi; however, no significant differences were found between control and all salinity treatments in L. macrosternum (post-hoc Scheffé: Control vs. 2 g/l P = 0.999; Control vs. 4 g/l P = 0.195; Control vs. 6 g/l P = 0.726) (Fig. 5)
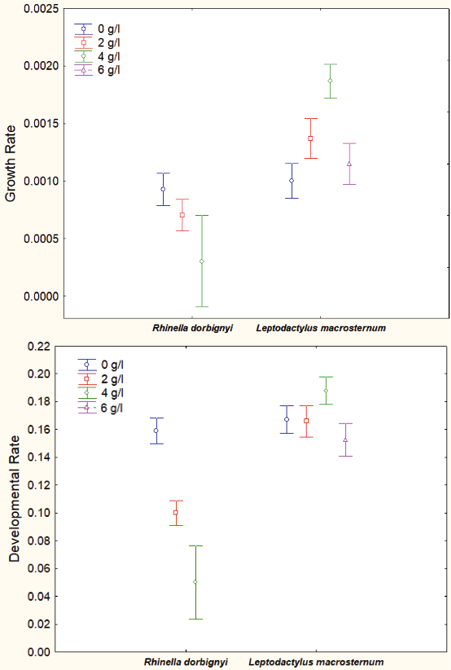
Figure 5. Growth and developmental rate at 20 days of exposure to different sublethal concentrations in Rhinella dorbignyi and Leptodactylus macrosternum.
Table 3. Univariate ANOVA F-values for growth rate and developmental rate of Rhinella dorbignyi and Leptodactylus macrosternum. P value: *P < 0.001
|
Source |
Growth rate |
Developmental rate |
||
|
df |
F |
df |
F |
|
|
R. dorbignyi |
||||
|
Treatment |
2 |
109.76* |
2 |
109.36* |
|
Time |
1 |
362.42* |
1 |
140.79* |
|
Treatment x time |
5 |
49.17* |
5 |
46.25* |
|
Error |
312 |
312 |
||
|
L. macrosternum |
||||
|
Treatment |
3 |
48.28* |
3 |
19.08* |
|
Time |
2 |
543.52* |
2 |
1685.49* |
|
Treatment x time |
6 |
19.23* |
6 |
9.91* |
|
Error |
368 |
368 |
||
DISCUSSION
Overall, our results demonstrate that salinity exerts a strong influence on survival, growth, and development in the early life stages of both studied species, although the magnitude and pattern of these effects differed between them. As predicted, increasing salinity levels led to higher mortality in both embryonic and larval stages, as reflected by the LC₅₀ estimates and survival analyses. In addition, sublethal salinity exposure negatively affected larval performance, particularly in Rhinella dorbignyi individuals evaluated in this study, where both body mass and developmental rate were significantly reduced under saline conditions. In contrast, Leptodactylus macrosternum individuals evaluated exhibited comparatively weaker responses, showing limited changes in survival, growth, and no significant differences in developmental rate across treatments. These findings are consistent with our initial objectives, as they reveal a clear relationship between salinity and mortality, while also highlighting species-specific differences in growth and developmental responses under sublethal conditions.
Salinity is widely recognized as a strong selective pressure on amphibian early life stages, primarily by imposing osmotic stress that disrupts ion balance and increases energetic costs associated with osmoregulation (Balinsky, 1981; Boutilier et al., 1992; Gomez-Mestre & Tejedo, 2003). In our study, increasing salinity produced a clear rise in embryonic and larval mortality in both species, with LC₅₀ values ranging approximately between 6 and 9.7 g/l in R. dorbignyi and between 7.1 and 8.8 g/l in L. macrosternum. Moreover, LC₅₀ values decreased with exposure time in R. dorbignyi, indicating a cumulative, time-dependent effect of salinity, whereas this trend was less evident in L. macrosternum. These results are consistent with previous studies showing that osmotic stress has both acute and chronic effects on amphibian survival (Sanzo & Hecnar, 2006; Karraker et al., 2008). In particular, Gomez-Mestre & Tejedo (2003) demonstrated that salinity reduces embryonic survival and that LC₅₀ values decline over time, reflecting the cumulative impact of prolonged exposure. Notably, these authors identified a tolerance threshold around 10–11 g/l, above which mortality becomes generalized across populations, a pattern that closely matches the upper tolerance limits observed in our study. Furthermore, although Gomez-Mestre & Tejedo (2003) found that populations from brackish environments exhibited higher initial tolerance, this advantage disappeared under sustained exposure, with all populations converging toward a similar lethal threshold. While our study did not explicitly address population-level adaptation, the interspecific differences observed here, particularly the more stable tolerance of L. macrosternum compared to R. dorbignyi, suggest that variation in osmotic tolerance may also occur at the species level. However, we consider that evaluating intraspecific variation, through the inclusion of multiple populations and families (see Gomez-Mestre & Tejedo, 2003), is essential to account for potential genetic variability underlying these responses. This is particularly relevant given that our study was based on a single clutch per species, which may have influenced the interspecific differences observed. Overall, these findings reinforce the idea that, despite differences in resistance and potential adaptive responses, amphibian early life stages share relatively conserved upper limits of salinity tolerance, beyond which survival is severely compromised.
Survival during the larval stage is a key component of fitness in amphibians, as small differences in early survival can scale up to significant effects on population dynamics (Semlitsch et al., 1988; Wells, 2007). In our sublethal experiment, we found that increasing salinity significantly reduced larval survival in both species, and that this effect was strongly modulated by exposure time. Mortality increased progressively over time, particularly in R. dorbignyi, where survival declined sharply at intermediate and high salinity treatments after 20 days of exposure. In contrast, L. macrosternum showed a comparatively weaker response, with smaller differences among treatments despite a general increase in mortality through time (Fig. 3). In parallel with these patterns, salinity also negatively affected key life-history traits. In R. dorbignyi, larval growth and developmental rate were both reduced under saline conditions, indicating a clear sublethal cost of osmotic stress. Conversely, in L. macrosternum, although some differences in body mass were detected, developmental rate did not differ significantly among treatments, suggesting a greater tolerance to salinity at sublethal levels. These results are consistent with previous studies showing that prolonged exposure to osmotic stress can amplify its negative effects, not only increasing mortality but also impairing growth and development through cumulative energetic costs associated with osmoregulation (Gomez-Mestre & Tejedo, 2003; Sanzo & Hecnar, 2006; Karraker et al., 2008). From an ecological perspective, the combined reduction in survival, growth, and developmental rate, particularly in R. dorbignyi, suggests that even moderate salinity levels, if persistent over time, can substantially reduce individual performance and delay development, potentially limiting recruitment and affecting long-term population persistence. This is especially relevant in temporary pond systems such as those studied here, where hydroperiod constraints may intensify the fitness consequences of slower growth and delayed development, reinforcing the importance of considering both the intensity and duration of salinity stress in ecological assessments.
In conclusion, the results suggest that, under the specific experimental conditions of this study, the L. macrosternum individuals evaluated exhibited differential tolerance behavior compared to R. dorbignyi specimens. Future multifactorial studies will be necessary to determine whether this trend persists at the population levels. The combined lethal and sublethal effects observed, particularly under prolonged exposure, highlight the importance of considering both the intensity and duration of osmotic stress. Interspecific differences in tolerance further indicate that species may respond differentially to increasing salinization, with potential consequences for community structure and population persistence (Semlitsch et al., 1988; Wells, 2007). These findings emphasize the need to monitor and regulate salinity levels in freshwater habitats, particularly in temporary pond systems, as even moderate increases may reduce recruitment and compromise population viability. Furthermore, our results underscore the need to extend this type of integrative approach to a wider range of Neotropical species, while also incorporating intraspecific variation through the evaluation of multiple populations and families, in order to account for potential genetic variability and improve predictions under ongoing environmental change.
CONFLICTS OF INTEREST
The authors declare no conflict of interest.
ACKNOWLEDGEMENTS
We thank students Arias, Agustina and Víctor, Paula for their invaluable assistance during fieldwork and laboratory sample processing. We also acknowledge the Cátedra de Ecología de Poblaciones, for providing a space for discussion and for supporting the implementation of these research activities as part of the students’ academic and scientific training. The authors have complied with all applicable Institutional Animal Careguide lines. The collecting permit was granted by Departamento de Biodiversidad de la Dirección de Recursos Naturales, Ministerio de Turismo, Corrientes province, Argentina, Disposciónes: 1114(17-09-2024) y 854(29-07-2025). This work was supported by Secretaría General de Ciencia y Técnica- Universidad Nacional del Nordeste (SGCyT-UNNE), PI-22Q001 granted to VG). We are very grateful for the continuous support of the Centro de Ecología Aplicada del Litoral (CECOAL-CONICET) and the Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste (FaCENA-UNNE).
AUTHOR CONTRIBUTIONS
FM developed the theoretical framework, conducted the fieldwork, and performed the statistical analyses. FM led the writing of the manuscript, with support from VG, who also contributed to the development of the theoretical framework, participated in the fieldwork, and assisted with laboratory experiments and the measurement of response variables.
REFERENCES
Albecker, M. A., & McCoy, M. W. (2017). Adaptive responses to salinity stress across multiple life stages in anuran amphibians. Frontiers in Zoology, 14: 40.
Balinsky, J. B. (1981). Adaptation of nitrogen metabolism to hyperosmotic environment. Journal of Experimental Zoology, 215: 335–350
Biggs, J., von Fumetti, S., & Kelly-Quinn, M. (2017). The importance of small waterbodies for biodiversity and ecosystem service: implications for policy makers. Hydrobiologia, 793: 3–39.
Boutilier, R. G., Stiffler, D. F., & Toews, D. P. (1992). Exchange of respiratory gases, ions, and water in amphibious and aquatic amphibians. In Environmental physiology of the amphibians. University of Chicago Press: 81–124.
Burraco, P., Torres-Montoro, J. C., & Gomez-Mestre, I. (2023). Larval plastic responses to warming and desiccation delay gonadal maturation in postmetamorphic spadefoot toads. Evolution, 17: 2687–2695.
Calhoun, A. J. K., Mushet, D. M., Bell, K. P., Boix, D., Fitzsimons, J. A., & Isselin-Nondeu, F. (2017). Temporary wetlands: challenges ans solutions to conserving a ‘disappearing’ ecosystem. Biological Conservation, 211: 3–11.
Cañedo-Argüelles, M., Kefford, B. J., Piscart, C., Prat, N., Schäfer, R. B., & Schulz, C. J. (2013). Salinisation of rivers: An urgent ecological issue. Environmental Pollution, 173: 157–167.
Carnevali, R. (1994). Fitogeografía de la provincia de Corrientes. Corrientes: gobierno de la Provincia de Corrientes, 324 p.
Chinathamby, K., Reina, R. D., Bailey, P. C. E., & Lees, B. K. (2006). Effects of salinity on the survival, growth and development of tadpoles of the brown tree frog, Litoria ewingii. Australian Journal of Zoology, 54: 97–105.
Chuang, M. F., Cheng, Y. J., Andersen, D. A., Borzée, C. S., Wu, Y. M., Chang, Y. J., Yang, Y., Jang, Y., & Kam, Y. C. (2022). Increasing salinity stress decreases the thermal tolerance of amphibian tadpoles in coastal areas of Taiwan. Scientific Reports, 12: 9014.
Dudgeon, D., Arthington, A. H., Gessner, M. O., Kawabata, Z. I., Knowler, D. J., Lévêque, C., Naiman, R. J., Prieur-Richard, A. H., Soto, D., Stiassny, M. L. J., & Sullivan, C. A. (2006). Freshwater biodiversity: Importance, threats, status and conservation challenges. Biological Reviews of the Cambridge Philosophical Society, 81: 163–182.
Gomez-Mestre, I., & Tejedo, M. (2003). Local adaptation of an anuran amphibian to osmotically stressful environments. Evolution, 57: 1889–1899.
Gomez-Mestre, I., Tejedo, M., Ramayo, E., & Estepa, J. (2004). Developmental alterations and osmoregulatory physiology of a larval anuran under osmotic stress. Physiological Biochemical Zoology, 77: 267–74.
Gosner, K. L. (1960). A simplified table for staging anurans embryos and larvae with notes of identification. Herpetologica, 16: 183–190.
Hall, E. M., Brunner, J. L., Hutzenbiler, B., & Crespi, E. J. (2020). Salinity stress increases the severity of ranavirus epidemics in amphibian populations. Proceedings Biological Science, 287: 20200062.
Herbert, E. R., Boon, P., Burgin, A. J., Neubauer, S. C., Franklin, R. B., Hopfensperger, K. N., Lamers, L. P. M., & Gell, P. (2015). A global perspective on wetland salinization: Ecological consequences of a growing threatto freshwater wetlands. Ecosphere, 6: 1–43.
Hintz, W. D., & Relyea, R. A. (2019). A review of the species, community, and ecosystem impacts of road salt salinisation in fresh waters. Freshwater Biology, 64: 1081–1097.
Hopkins, G. R., & Brodie, E. D., Jr. (2015). Occurrence of amphibians in saline habitats: A review and evolutionary perspective. Herpetological Monographs, 29: 1–27.
Karraker, N. E., Gibbs, J. P., & Vonesh, J. R. (2008). Impacts of road deicing salt on the demography of vernal pool-breeding amphibians. Ecological Applications, 18: 724–734.
Kaushal, S. S., Groffman, P. M., Likens, G. E., Belt, K. T., Stack, W. P., Kelly, V. R., Band, L. E., & Fisher, G. T. (2005). Increased salinization of fresh water in the northeastern United States. Proceedings of the National Academy of Sciences USA, 102: 13517–13520.
Relyea, R. A., Schermerhorn, C. X., Mattes, B. M., & Shepard, I. D. (2023). Phenotypically plastic responses to freshwater salinization in larval amphibians: Induced tolerance and induced sensitivity. Environmental Pollution, 337: 122588.
Relyea, R. A., Mattes, B. M., Schermerhorn, C. X., & Shepard, I. D. (2024). Freshwater salinization and the evolved tolerance of amphibians. Ecology and Evolution, 14: e11069.
Sanabria, E., Quiroga, L., Vergara, C., Banchig, M., Rodriguez, C., & Ontivero, E. (2018). Effect of salinity on locomotor performance and thermal extremes of metamorphic Andean Toads (Rhinella spinulosa) from Monte Desert, Argentina. Journal of Thermal Biology, 74: 195–200.
Sanzo, D., & Hecnar, S. J. (2006). Effects of road salt (NaCL) on larval wood frogs (Rana sylvatica). Environmental Pollution, 140: 247–256.
Semlitsch, R. D., Scott, D. E., & Pechmann, J. H. K. (1988). Time and size at metamorphosis related to adult fitness in Ambystoma talpoideum. Ecology, 68: 184–1992.
Sokal, R.R. & Rohlf, F.J. (1995) Biometry: The Principles and Practice of Statistics in Biological Research. 3rd Edition, W.H. Freeman and Co., New York.
Sparks, T. 2000. Statistics in ecotoxicology. John Wiley and Sons, Chichester, U.K.
Stănescu, F., Székely, D., Székely, P., Topliceanu, S. & Cogalniceanu, D. (2017).The impact of salinity on early developmental stages in two sympatric spadefoot toads and implications for amphibian conservation in coastal areas Hydrobiologia 792: 357–366 .
StatSoft, Inc. (2007). STATISTICA (data analysis software system), version 8.0. www.statsoft.com.
Székely, D., Cogalniceanu, D., Székely, P., Armijos-Ojeda, D., Espinosa-Mogrovejo, V., & Denoël, M. (2020). How to recover from a bad start: size at metamorphosis affects growth and survival in a tropical amphibian. BMC Ecology, 20: 24
Vörösmarty, C. J., McIntyre, P. B., Gessner, M. O., Dudgeon, D., Prusevich, A., Green, P., Glidden, S., Bunn, S. E., Sullivan, C. A., Liermann, C. R., & Davies, P. M. (2010). Global threats to human water security and river biodiversity. Nature, 467: 555–561. Erratum in: Nature. 2010 Nov 11; 468(7321): 334.
Wells, K. D. (2007). The ecology and behavior of amphibians. University of Chicago Press.
Wu, C. S., & Kam, Y. C. (2009). Effects of salinity on the survival, growth, development, and metamorphosis of Fejervarya limnocharis tadpoles living in brackish water. Zoological Science, 26: 476–482.